不同恢复模式下干热河谷幼龄印楝和大叶相思生物量及其分配
2012-05-30高成杰唐国勇孙永玉张春华谢青海
高成杰, 唐国勇,, 孙永玉,, 张春华,,谢青海, 李 昆,
(1.中国林业科学研究院 资源昆虫研究所,云南 昆明 650224;2.国家林业局 云南元谋荒漠生态系统定位研究站,云南 昆明 650224)
生物量是研究植被生产力的基础和估算植被碳库的关键参数,也是衡量林地生产力和林地经营以及植被恢复效果的重要指标。生物量分配的研究是分析植物碳分配的基础和植物结构功能最有效的工具,对研究生态系统碳循环和营养物质的分配具有重要意义[1-4]。20世纪60年代以来,植被生物量和生产力从立地到区域和全球尺度上得到了广泛的观测、分析和模拟[5-6]。近年来,有大量研究报道了不同胁迫或干扰下植物生物量分配策略及其形成机制[7-8]。在中国某些造林困难区(如干热河谷)树种生物量分配的研究还较为鲜见。西南干热河谷是中国特有的生态脆弱区,由于植被破坏和水土流失严重,加速了土壤退化[9]。为了改善生态环境,当地政府依托长江上中游水土保持重点防治工程(长治)及防护林体系建设工程(长防)等项目引进了众多生态适应性强的多功能树种,并营造了各种类型的人工植被,如印楝Azadirachta indica和大叶相思Acacia auriculiformis人工林等。印楝是生产生物农药的关键原料,已成为中国荒漠化地区水土保持和植被恢复的主要树种[10];而大叶相思因具根瘤,是良好的辅佐、护土、改土树种[11]。目前,对于干热河谷地区引进树种生物量方面的研究已有报道,如杨忠等[12]对元谋干热河谷不同坡地类型下桉树Eucalyptus spp.人工林生物量进行了研究。刘方炎等[13]从植株生长状况和生物量等方面,将该地区几种引进树种和当地典型的天然次生植被进行了比较。然而,在干热河谷人工林营造过程中,不同种植模式下人工林生物量及其分配还缺乏对比研究。对干热河谷人工林生物量及其分配特征进行研究,有助于了解该地区人工林生态系统中的营养积累与分布特征、树种混交效益和竞争关系。基于此,以元谋干热河谷9年生印楝和大叶相思为研究对象,对印楝纯林、大叶相思纯林及印楝与大叶相思混交林内树种和林分生物量及其分配进行对比研究,拟建立该地区印楝和大叶相思生物量回归模型,探明其各器官生物量的异速生长关系,旨在为中国干热河谷造林困难区人工植被恢复和经营提供理论基础。
1 研究区概况
1.1 地理位置和自然条件
试验点位于国家林业局云南元谋荒漠生态系统定位研究站(25°40′N,101°52′E)。该点海拔为1120 m,年平均气温 21.9℃,最高月平均气温27.1℃ (5月),最低月平均气温14.5℃ (12月),年降水量634.0mm,其中90%以上的降水集中在6-10月;年蒸发量为3911.2mm,年相对湿度53%,年均干燥度2.8,≥10℃的年积温8003℃,日照时数 2670 h,无霜期350~365 d,属南亚热带干热季风气候区。植被以坡柳Dodonaea viscose,余甘子Phyllanthus emblica,扭黄茅Heteropogon contortus和三芒草Aristida adscensionis等为主,土壤类型为燥红土[14]。
1.2 试验林营建
在研究站内选取约2.2 hm2的退耕地(退耕前耕地主要种植玉米Zea mays等)为试验地。试验地内立地条件相对一致,坡度为7°~12°,以阳坡为主。于2001年5月(雨季初期)选择印楝和大叶相思等树种,按照2 m×3 m株行距和70 cm×60 cm×60 cm植塘规格进行容器苗造林,并进行封禁管理。
2 研究方法
2.1 样地调查与生物量测定
于2010年11月上旬(雨季末期)分别布设9年生印楝纯林、大叶相思纯林及印楝与大叶相思混交林的典型样地各2块,样地面积为600 m2(20 m×30 m)。生物量测定采用间接收获法,即按平均标准木和林分密度估测林分生物量[15]。对样地内林木进行每木检尺,计算出林木的平均胸径、平均树高和平均冠幅等测树指标,并采样测定样地内土壤主要理化性质(表1)。根据每木检尺结果,在每种林分内分树种选出标准木各6株,共24株,用于单株生物量和林分生物量的测定。地上部分采用分层切割法并按0.5 m区分段截取圆盘,根系采用全挖法[15];分别测定各器官(干、皮、枝、叶和根)的鲜质量,同时随机抽取各器官样品约500 g在85℃下烘干至恒量,计算各器官的干质量。
2.2 数据处理和分析方法
通过单因素方差分析比较同一树种单株生物量或器官生物量在不同林分下差异显著性。按不同径级(表1)选取印楝共17株(纯林内10株,混交林内7株,不同林分树种各个径级1~2株),大叶相思共16株(纯林内11株,混交林内5株),分别建立纯林和混交林下印楝和大叶相思各器官之间的异速生长方程,以及该地区幼龄印楝和大叶相思生物量与测树因子(D或D2H)的回归方程。植物各器官生物量异速生长方程通常采用幂函数(y=axb)的形式表示[16-18]。其中,y与x为不同器官生物量;幂指数b为各器官间的异速生长系数。当b=1时,这种异速生长即表现为等速生长[19]。方差分析和各器官生物量异速生长方程的建立在SPSS 13.0上完成,并利用该软件的Curve Estimation分别对生物量与胸径(D)和胸径平方乘以树高(D2H)进行拟合,建立生物量估测回归方程,选出拟合效果较好、关系密切的模型作为生物量估算模型。
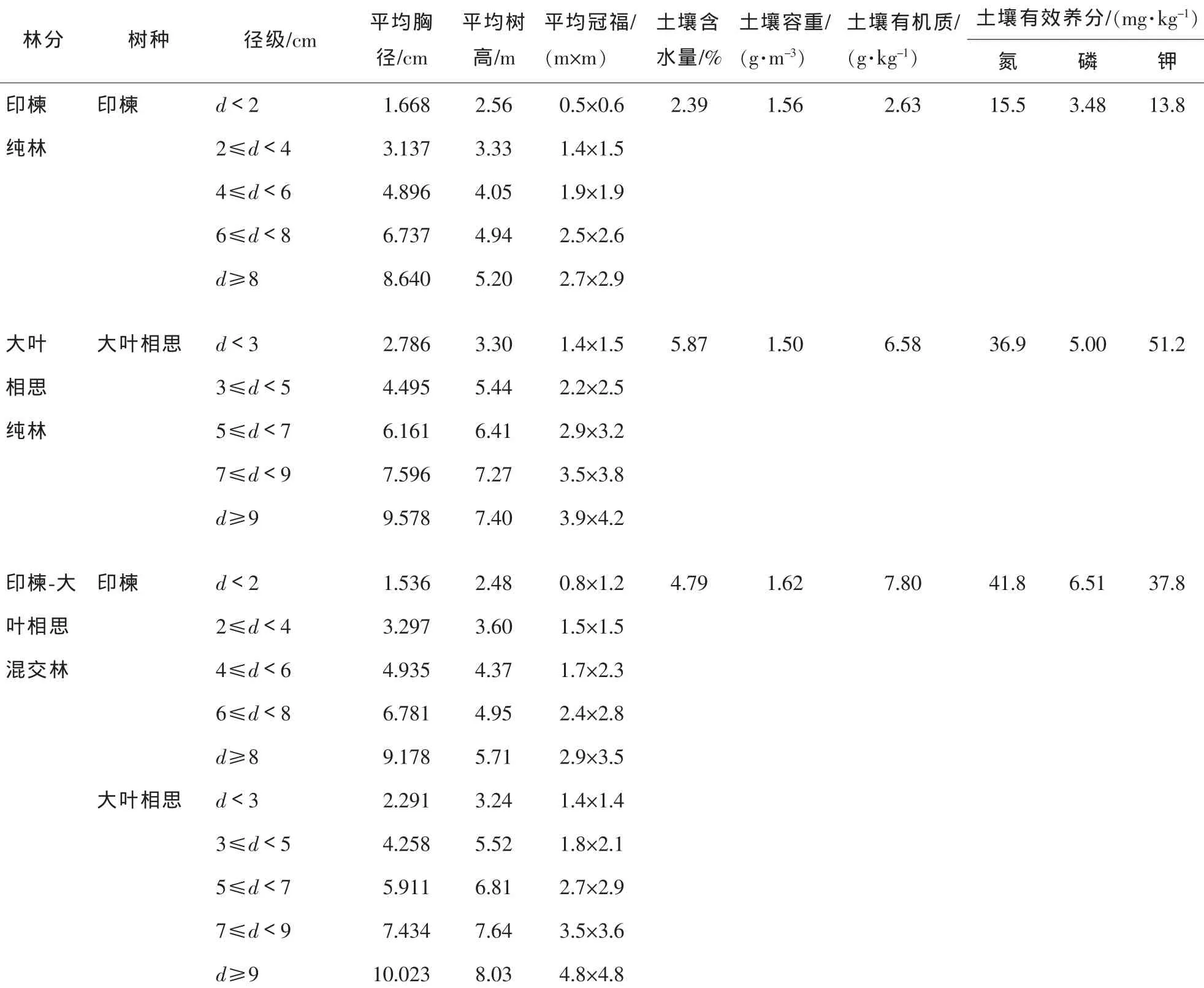
表1 印楝和大叶相思林基本特征Table1 Basic characteristics of Azadirachta indica and Acacia auriculiformis stands
3 结果与分析
3.1 单株生物量与林分生物量及其分配
基于印楝和大叶相思人工林样地调查资料,运用平均标准木法对树种各器官的生物量和林分生物量进行推算(表2)。结果表明,印楝与大叶相思混交种植后,混交林内印楝单株生物量[(5.713±0.663)kg·株-1]较纯林印楝[(4.898 ± 0.722) kg·株-1]提高了 16.6%; 而大叶相思[(14.943 ± 1.975) kg·株-1]较纯林大叶相思[(17.377±2.067)kg·株-1]下降了14.0%,但同一树种在不同林分下单株生物量及器官生物量差异均不显著(P>0.05)。混交林内印楝地上(干、皮、枝、叶)生物量较纯林印楝提高了13.2%~69.3%,尤其是叶(69.3%);而地下(根)生物量下降了7.0%。混交林内大叶相思干、叶和根较纯林大叶相思分别提高了5.5%,17.7%和4.6%;而皮和枝分别下降了28.1%和36.0%。纯林和混交林下,印楝和大叶相思地下部分与地上部分生物量之比(根冠比)不同,混交林印楝根冠比(0.280±0.032)较纯林(0.400±0.090)小(P<0.05), 而混交林大叶相思(0.163 ± 0.007)较纯林(0.132 ± 0.016)大(P>0.05)。
从各器官生物量分配比例看(表2),同一树种在纯林和混交林下分配格局不同。在纯林和混交林下,印楝各器官生物量大小顺序分别为干>根>枝>皮>叶和干>枝>根>叶>皮;而大叶相思分别为枝>干>根>叶>皮和干>枝>根>叶>皮。干热河谷印楝和大叶相思干生物量分配比例为31.3%~38.2%,且以混交林树种高于纯林树种。枝(21.9%~43.3%)和根(11.4%~27.2%)的分配比例较高,尤其是纯林大叶相思,枝的分配比例(43.3%)要高于干(31.2%)。印楝枝和叶的分配比例以混交林高于纯林,皮和根以纯林高于混交林;大叶相思叶和根的分配比例以混交林高于纯林,皮和枝以纯林高于混交林。
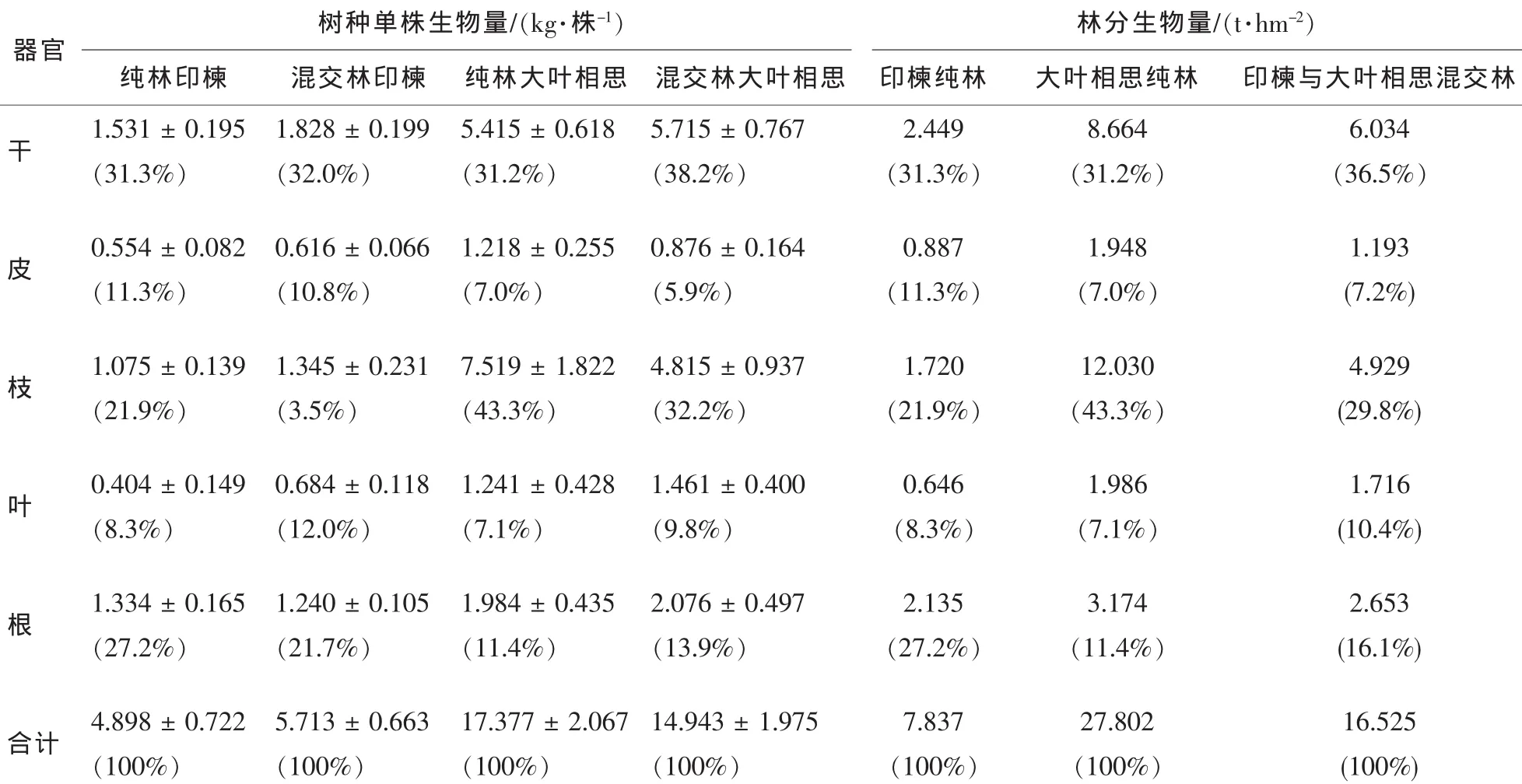
表2 林木与林分生物量及其分配Table2 Biomass and its allocation of plants and stands
从林分生物量来看(表 2), 印楝与大叶相思混交林生物量(16.525 t·hm-2)介于印楝纯林(7.837 t·hm-2)和大叶相思纯林(27.802·hm-2)之间,是印楝纯林的2.1倍,而较大叶相思纯林下降了40.6%。混交林各器官生物量分配比例为干>枝>根>叶>皮,其中,干和叶的分配比例高于纯林,而皮、枝和根的分配比例均居于印楝纯林和大叶相思纯林之间。
3.2 生物量回归模型
3.2.1 生物量与测树因子的回归模型 依据植物相对生长规律,以及标准木的干、枝、叶、根、地上和整株生物量(W)与胸径(D),树高(H)的实测数据,分别对生物量与测树因子(D或D2H)进行拟合,结果以幂函数曲线(W=axb)拟合效果最佳(表3)。经F检验表明,相关性均达到显著水平(P<0.001)。除大叶相思叶生物量与测树因子(D或D2H)拟合模型的相关系数为0.793和0.762,各器官、地上生物量和整株生物量数学模型的相关系数均超过0.9。
回归模型W=axb的幂指数反映了生物量与测树因子(D或D2H)的生长关系。以胸径(D)建立的回归模型的幂指数为1.515~3.544,均大于1。表明印楝和大叶相思胸径与各器官、地上生物量和整株生物量呈异速生长关系,且胸径的异速生长速率要落后于各器官。以D2H建立的回归模型以枝生物量拟合的幂指数大于1,其他各器官、地上生物量和整株生物量拟合的幂指数为0.569~0.881,均小于1;表明印楝和大叶相思胸径树高(D2H)的异速生长速率高于其他各器官,但落后于枝。
3.2.2 树种器官异速生长关系 印楝和大叶相思各器官的异速生长关系均呈显著或极显著水平,相关系数为0.669~0.999(表4)。其中纯林印楝和混交林印楝叶与干均近于等速生长(幂指数分别为1.040和0.952),其他各器官之间呈较明显的异速生长关系(幂指数为0.475~0.906,1.224~1.855),且纯林和混交林下印楝各器官异速生长速率表现为枝>叶/干>根,地上部分>地下部分。不同林分下大叶相思根与干、根与地上部分异速生长关系不同,纯林大叶相思根的异速生长速率落后于干和地上部分,而混交林内则近于等速生长(幂指数分别为1.069和1.010)。其他各器官之间呈较明显的异速生长关系(幂指数为0.474~0.821,1.191~1.358),且纯林和混交林下大叶相思各器官异速生长速率表现为枝>干/根>叶,地上部分>地下部分。
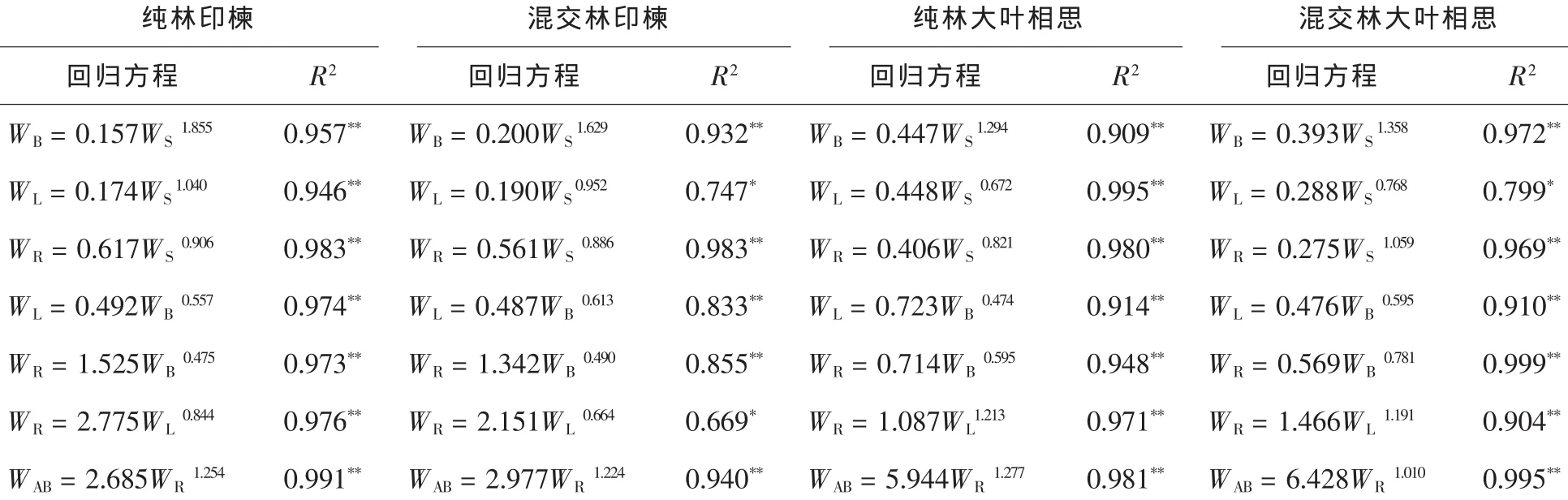
表4 印楝与大叶相思各器官异速生长关系Table4 Allometry relationship of organs of A.indica and A.auriculiformis in dry-hot valley
4 结论与讨论
人工林生物量与生物因素和非生物因素密切相关,如地理位置和气候条件、林分内的水热条件、土壤条件以及林分类型、年龄和林分密度等[16,20-21]。干热河谷地区降水量小,蒸发量大,水热矛盾突出,植物生长环境受到极大破坏,乡土树种很难正常更新繁殖;而通过引进树种按照纯林和混交林等不同恢复模式营造的人工林在水土保持、土壤改良、生态防护等方面发挥了重要作用[13]。在相同种植密度下,干热河谷9年生印楝和大叶相思在纯林和混交林下生物量不同。将印楝和大叶相思混交种植后,提高了印楝生物量,而大叶相思生物量却有所下降。许多研究结果表明,通过营造混交林能明显提高林地养分水平,改善林地养分状况,从而提高林地生产力和林分稳定性[20-22]。大叶相思因其具有根瘤,可在一定程度上改善林地土壤理化性状,提高土壤肥力。本研究中,混交林内土壤有机质和有效养分(氮、磷、钾)质量分数均高于印楝纯林(表1),这可能是造成纯林印楝生物量低于混交林印楝的主要因素。而纯林和混交林下大叶相思生物量的差异可能与大叶相思种内和种间对水分和养分的竞争有关。这有待于进一步通过系统的栽培试验加以验证。
植物生物量分配受环境可利用资源(如光照、水分和养分等)、物种和植株大小等变化的影响[16,23]。植物在生长发育过程中,当某些环境因子成为影响植物生长发育的限制性因素时,为了保证其正常生长不受制约,不断地调整生长和营养物质的分配以及改变根冠比等形态特征指标来适应环境变化[23]。Huston等[24]和Tilman[25]认为,植物地下部分与地上部分生物量的大小反映了植物对土壤养分或光照的需求和竞争能力。根冠比大,反映出对养分的需求和竞争能力强;反之,则反映出对光照的需求和竞争能力强[26-27]。干热河谷混交林印楝根冠比纯林小,反映了纯林印楝对养分的需求高于混交林,混交林内印楝冠幅和树高都不及大叶相思(表1),对光照的需求则更为强烈;而混交林内大叶相思根冠比较纯林大,其对养分的需求和竞争可能高于纯林。与中国大多数人工林树种干生物量分配比例相比,如日本落叶松Larix kaempferi(51.3% ~ 64.3%)[28], 马尾松 Pinus massoniana(51.2% ~ 63.3%)[29], 橡胶 Hevea brasiliensis(52.4% ~ 67.3%)[4], 华北落叶松 Larix principis-rupprechtii(46.3% ~ 62.6%)[30]等, 干热河谷印楝和大叶相思干生物量分配比例较低(31.2%~38.2%),枝、叶和根则相对较高,这可能与干热河谷水分缺乏,养分贫瘠的环境有关,反映了林木在水分和养分胁迫下通过自身生理过程适应环境的一种策略[27,31-32]。
国内外研究者[31,33-34]利用多种生物量模型(如二次曲线、三次曲线和幂函数曲线)来估计林木的生物量,尤以幂函数曲线最常用。其变量主要有胸径(D),胸径平方(D2),树高(H),胸径平方乘以树高(D2H),也有的模型加上树龄、树冠等变量[33]。但在森林群落调查中,胸径的测量较准确,树高的测量通常存在较大的误差[35]。研究表明,生物量和胸径之间存在着很好的相关关系,以胸径为单变量的生物量模型在多种森林生物量预测中广泛使用[4,35-36]。本研究显示,以胸径为单变量建立印楝和大叶相思生物量数学模型也具有较高的相关性,各器官及总生物量数学模型的相关系数均达到极显著水平(P<0.01)。另外,由于林木林龄相同(均为9年生),印楝或大叶相思树高差异不大,对树高的测量相对较为准确,以D2H为自变量建立的数学模型相关系数也较高。但利用相对生长方程估测林木生物量时,以胸径(D)作为单一变量的生物量回归模型更具实用性。
异速生长广泛存在于植物中[37-38],植物的异速生长与生物量分配、植物对资源的利用、对异质环境的形态适应有密切的关系[16-17]。通过异速生长方程来检验生物量分配与个体大小的关系,能够较好地解释物种和个体大小对生物量分配的影响。干热河谷印楝和大叶相思生物量与测树因子(D或D2H)呈异速生长关系,且胸径(D)异速生长速率落后于树种生物量;而D2H异速生长速率则高于树种生物量(大叶相思枝除外)。树种各器官之间也均呈异速生长关系,且同一树种各器官在不同林分下异速生长关系差异并不大。纯林和混交林下印楝各器官异速生长速率表现为枝>叶/干>根,地上部分>地下部分;而大叶相思表现为枝>干/根>叶,地上部分>地下部分。异速生长关系是植物生理和形态共同作用的结果[17],因此,与植物异速生长关系相关的生理学、生态学等内在机制方面的探讨还有待深入。
[1]OVERMAN J P M, WITTE H J L, SALDARRIAGA J G.Evaluation of regression models for above-ground biomass determination in Amazon rainforest[J].J Trop Ecol, 1994, 10: 207-218.
[2]樊后保,李燕燕,苏兵强,等.马尾松-阔叶树混交异龄林生物量与生产力分配格局[J].生态学报,2006,26(8): 2463-2473.FAN Houbao, LI Yanyan, SU Bingqiang, et al.Allocation pattern of biomass and productivity in the mixed unevenaged stands of masson’s pine and hardwood species [J].Acta Ecol Sin, 2006, 26 (8): 2463-2473.
[3]GARKOTI S C.Estimates of biomass and primary productivity in a high-altitude maple forest of the west central Himalayas [J].Ecol Res, 23: 41-49.
[4]唐建维,庞家平,陈明勇,等.西双版纳橡胶林的生物量及其模型[J].生态学杂志,2009,28(10):1942-1948.TANG Jianwei, PANG Jiaping, CHEN Mingyong, et al.Biomass and its estimation model of rubber plantations in Xishuangbanna, Southwest China [J].China J Econol, 2009, 28 (10): 1942-1948.
[5]DIXON R K, SOLOMON A M, BROWN S, et al.Carbon pools and flux of global forest ecosystems[J].Science, 1994,263:185-190.
[6]SCURLOCK J M O, CRAMER W, OLSON R J, et al.Terrestrial NPP: towards a consistent data set for global model evaluation [J].Ecol Appl, 1999, 9 (3): 913-919.
[7]CAO Kunfang, OHKUBO T.Allometry, root/shoot ratio and root architecture in understory saplings of deciduous dicotyledonous trees in central Japan [J].Ecol Res, 1998, 13 (2): 217-227.
[8]赵彬彬,牛克昌,杜国桢.放牧对青藏高原东源高寒草甸群落27种植物地上生物量分配的影响[J].生态学报,2009, 29 (3): 1596-1606.ZHAO Binbin, NIU Kechang, DU Guozhen.The effect of grazing on above-ground biomass allocation of 27 plant species in an alpine meadow plant community in Qinghai-Tibetan Plateau [J].Acta Ecol Sin, 2009, 29 (3): 1596-1606.
[9]张建平,张信宝,杨忠,等.云南元谋干热河谷生态环境退化及恢复重建试验研究[J].西南师范大学学报:自然科学版, 2001, 26(6): 733-739.ZHANG Jianping, ZHANG Xinbao, YANG Zhong, et al.Studies on eco-environment degradation and the experiment of environment recovering and rebuilding [J].J Southwest China Norm Univ Nat Sci Ed, 2001, 26 (6): 733-739.
[10]张燕平,赖永琪,彭兴民,等.印楝的世界地理分布与引种栽培概况[J],林业调查规划,2002,27(3):98-101.ZHANG Yanping, LAI Yongqi, PENG Xingming, et al.Global distribution and introduction cultivation state of Azadirachta indica [J].For Inventory&Plann, 2002, 27 (3): 98-101.
[11]许松葵,王相娥,谢腾芳,等.不同密度大叶相思幼林的土壤肥力[J].华南农业大学学报,2008,29(2):79-81.XU Songkui, WANG Xiang’e, XIE Tengfang, et al.Soil fertility of young Acacia auriculiformis stands with different densities [J].J South China Agric Univ, 2008, 29 (2): 79-81.
[12]杨忠,张建辉,徐建忠,等.元谋干热河谷不同岩土组成坡地桉树人工林生长特征初步研究[J].水土保持学报, 2000, 14 (5): 1-6, 34.YANG Zhong, ZHANG Jianhui, XU Jianzhong, et al.Growth responses of Eucalyptus camaldulensis Dehnl artificial population to slopes in arid-hot valleys, Yuanmou, Yunnan [J].J Soil Water Conserv, 2000, 14 (5): 1-6, 34.
[13]刘方炎,李昆,马姜明.金沙江干热河谷几种引进树种人工植被的生态学研究[J].长江流域资源与环境,2008,17(3): 468-474.LIU Fangyan, LI Kun, MA Jiangming.Ecological study on several man-made forests of introduced species in Jinsha-jiang dry-hot valley [J].Resour Environ Yangtze Basin, 2008, 17 (3): 468-474.
[14]李昆,曾觉民,赵虹.金沙江干热河谷造林树种游离脯氨酸含量与抗旱性关系[J].林业科学研究,1999,12(1):103-107.LI Kun, ZENG Juemin, ZHAO Hong.Drought resistance in relation to free proline in the major tree species in dry and hot river valley of Jin Sha [J].For Res, 1999, 12 (1): 103-107.
[15]董鸣.陆地生物群落调查观测与分析[M].北京:中国标准出版社,1997:62-64.
[16]ENQUIST B J, NIKLAS K J.Global allocation rules for patterns of biomass partitioning in seed plants [J].Science,2002, 295: 1517-1520.
[17]MCCARTHY M C, ENQUIST B J.Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation [J].Funct Ecol, 2007, 21: 713-720.
[18]WERNER P A, MURPHY P G.Size-specific biomass allocation and water content of above-and belowground components of three Eucalyptus species in a northern Australian savanna [J].Aust J Bot, 2001, 49: 155-167.
[19]GOULD S J.Allometry and size in ontogeny and phylogeny [J].Biol Rev, 1966, 41: 587-640.
[20]刘芳.杉木光皮桦纯林及混交林生物量[J].浙江林学院学报,2002,19(2):143-147.LIU Fang.Growth and biomass of pure stands of Betula luminifera and Cunninghamia lanceolata and their mixed forest[J].J Zhejiang For Coll, 2002, 19 (2): 143-147.
[21]安慧,上官周平.密度对刺槐幼苗生物量及异速生长模式的影响[J].林业科学,2008,44(3):151-155.AN Hui,SHANGGUAN Zhouping.Effects of density on biomass and allometric pattern of Robinia pseudoacacia seedling [J].Sci Silv Sin, 2008, 44 (3): 151-155.
[22]贾黎明.固氮树种与非固氮树种混交林研究现状[J].世界林业研究,1998,11(1):20-26.JIA Liming.The review of mixtures of nitrogen-fixing and non-nitrogen-fixing tree species [J].World For Res, 1998, 11(1): 20-26.
[23]POORTER H, NAGEL O.The role of biomass allocation in the growth response of plants to different levels of light,CO2, nutrients and water: a quantitative review [J].Aust J Plant Physiol, 2000, 27: 595-607.
[24]HUSTON M, SMITH T.Plant succession: life history and competition [J].Am Nat, 1987, 130: 168-198.
[25]TILMAN D.Plant Strategies and the Dynamics and Structure of Plant Communities [M].Princeton: Princeton University Press, 1988: 52-97.
[26]王军邦,王政权,胡秉民.不同栽植方式下紫椴幼苗生物量分配及资源利用分析[J].植物生态学报,2002,26(2): 677-683.WANG Junbang, WANG Zhengquan, HU Bingmin.Biomass allocation and resource use of Tilia amurensis juvenile under different planting treatment[J].Acta Pytoecol Sin, 2002, 26 (2): 677-683.
[27]吴楚,王政权,范志强,等.氮胁迫对水曲柳幼苗养分吸收、利用和生物量分配的影响[J].应用生态学报,2004, 15 (11): 2034-2038.WU Chu, WANG Zhengquan, FAN Zhiqiang, et al.Effects of nitrogen stress on nutrient uptake by Fraxinus mandshurica seedlings and their biomass distribution [J].Chin J Appl Ecol, 2004, 15 (11): 2034-2038.
[28]王永祥,吴耀先,李长斌,等.日本落叶松人工林生物量及器官分配规律的研究[J].辽宁林业科技,1999(2):17-20.WANG Yongxiang, WU Yaoxian, LI Changbin, et al.Study on biomass of Larix kaempferi plantation and its distribution [J].J Liaoning For Sci Technol, 1999 (2): 17-20.
[29]蔡秀珠.人为干扰强度对马尾松人工林生物量及其分配的影响[J].中南林学院学报,2006,26(2):51-54.CAI Xiuzhu.Effect of different disturbances on biomass and its distribution of Pinus massoniana plantation [J].J Cent South For Univ, 2006, 26 (2): 51-54.
[30]罗云建,张小全,王效科,等.华北落叶松人工林生物量及其分配模式[J].北京林业大学学报,2009,31(1):13-18.LUO Yunjian, ZHANG Xiaoquan, WANG Xiaoke, et al.Biomass and its distribution pattern of Larix principis-rupprechtii plantations in northern China [J].J Beijing For Univ, 2009, 31 (1): 13-18.
[31]肖冬梅,王森,姬兰柱.水分胁迫对长白山阔叶红松林主要树种生长及生物量分配的影响[J].生态学杂志,2004, 23 (5): 93-97.XIAO Dongmei, WANG Sen, JI Lanzhu.Influence of water stress on growth and biomass allocation of dominant tree species in mixed forest of broad-leaved and Korean pine at Changbai Mountain [J].Chin J Ecol, 2004, 23 (5): 93-97.
[32]贺海波,李彦.干旱、盐胁迫条件下2种盐生植物生物量分配对策的研究[J].干旱区研究,2008,25(2):242-247.HE Haibo,LI Yan.Study on measures of biomass allocation of two desert halophyte species under drought and salt stress [J].Arid Zone Res, 2008, 25 (2): 242-247.
[33]黎燕琼,郑绍伟,宿以明,等.岷江上游干旱河谷引种番麻的生态适应性及生物量预测模型[J].生态学报,2009, 29 (9): 4820-4826.LI Yanqiong, ZHENG Shaowei, SU Yiming, et al.Growth and allometric biomass equation of introduced Agave americana L.in upper reaches of Minjiang River [J].Acta Ecol Sin, 2009, 29 (9): 4820-4826.
[34]KAJIMOTO T, MATSUURA Y, OSAWA A, et al.Size-mass allometry and biomass allocation of two larch species growing on the continuous permafrost region in Siberia [J].For Ecol Manage, 2006, 222: 314-325.
[35]CHAVE J, RIERA B, DUBOIS M A.Estimation of biomass in a neotropical forest of French Guiana Spatial and temporal variability [J].J Trop Ecol, 2001, 17: 79-96.
[36]SEGURA M, KANNINEN M.Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica [J].Biotropica, 2005, 37: 2-8.
[37]LIU Rentao, BI Runcheng, ZHAO Halin.Biomass partitioning and water content relationships at the branch and whole-plant levels and as a function of plant size in Elaeagnus mollis populations in Shanxi, North China [J].Acta Ecol Sin, 2009, 29: 139-143.
[38]WEINER J.Allocation, plasticity and allometry in plants [J].Perspect Plant Ecol Evol Syst, 2004, 6 (4): 207-215.
