贵州喀斯特区野生葛藤群落主要种群生态位
2012-05-30李安定李苇洁谢元贵龙秀琴
李安定,李苇洁,彭 熙,谢元贵,龙秀琴
(贵州科学院喀斯特资源环境与发展研究中心,贵州 贵阳 550001)
生态位一词首先由Grinnel引入到生态学领域中,并把它定义为物种在环境中的最后分布单位[1-3]。生态位与种间竞争有关,同资源的利用密不可分[4]。生态位研究是近代生态学理论的重要内容,种的生态位宽度和种间生态位重叠被认为是物种多样性及群落结构的决定因素,反映了种群对资源的利用能力[5-6]及其群落的稳定性。生态位宽度和生态位重叠的计算分析对不同植物环境适应性的比较具有重要作用[7]。葛藤Pueraria lobata是豆科Leguminosae葛属Pueraria植物。葛藤在中国的分布极广,南起广东、广西,北到东北各省都有生长,而以东南和西南各省区盛产。在国外,俄罗斯、朝鲜、日本、越南、印度、马来西亚等地也有分布[8-9]。美国曾从中国引种栽培繁殖。葛藤喜光照充足、温暖潮湿,土壤肥沃的环境,但同时也耐寒、耐旱、耐瘠薄,抗病虫害。对土质要求不严,在微酸性的红壤、黄壤、花岗岩砾土、砂砾土及中性泥砂土、紫色土均可生长,也能生长于石缝、荒坡、砾石地及喀斯特石质山上。其根茎可深入地下3 m以上,具有很强的抗旱和固土能力,加上匍匐于地面的多枝长蔓,对保持水土有着重要的作用。葛藤生长要求的年降水量为400mm以上,适宜的降水量约为1000mm,夏季高温多雨,对葛藤的生长最为有利[10]。葛藤的根、茎、花、叶及果均可利用,是一种药食同源植物,同时具有较好的饲用开发价值。近年来,研究珍稀濒危植物的种群生态位特征,探讨濒危植物对空间资源的利用及其群落结构等方面的研究报道逐渐增多[11-12],对濒危种群保护措施的制定和实施非常重要。通过对喀斯特地区葛藤群落组成类型中16个优势种进行研究,提示葛藤在喀斯特石漠化区的功能地位、生态适应性及生态相似性,探讨种群对资源的利用状况和发展趋势,从而为合理开发利用葛藤及有效治理石漠化提供科学依据。
1 研究区概况
贵州省清镇市卫城镇的阳山村和羊皮洞位于黔中典型的喀斯特高原盆地中心区,地处26°49′20″~26°51′43″N, 106°19′30″~106°19′37″E, 地貌主要为低山丘陵、 洼地, 洼地面积比例较大, 以黄壤、 黄色石灰土为主;海拔1239~1203 m,其中4-8月降水量占全年降水量的77.3%,年蒸发量700mm左右,无霜期278 d。该区气候属亚热带季风区,平均气温为14.0℃,年降水量为1192.5mm,≥10℃的活动积温为4500℃,最热月(7月)均温为22.0℃,最冷月(1月)均温3.6℃;年辐射总量为3.59×106kJ·m-2;冬无严寒夏无酷暑。
2 研究方法
2.1 取样及数据处理
2010年6-8月,对清镇市阳山村石水沟石漠化区葛藤的分布环境特点进行全面考察,发现葛藤主要分布在石漠化地区早期到中期阶段,即群落演替的草本群落阶段、灌草群落阶段。岩石裸露率高达60%以上。 采取典型取样法(表 1), 设置 10 m × 10 m 的 9 个样地,26°51′64″N,106°19′30″E,海拔为1239 m,并记录物种组成(类型)、高度、盖度、株数、地径等。综合各物种在样地中的盖度与出现频率,共选取16个优势种,进行生态位宽度及重叠分析。
2.2 生态位测试方法
重要值的计算:采用重要值作为反映植物与环境关系的重要指标。计算公式如下:植物重要值=(相对盖度+相对多度+相对频度)/300。
生态位宽采用Shannon-Wiener生态位宽度指数计算[13-14]:

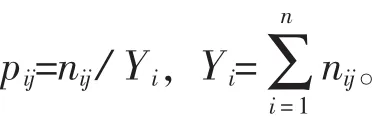
生态位相似性比例是指2个物种利用资源的相似程度,其计算公式[13-14]为:
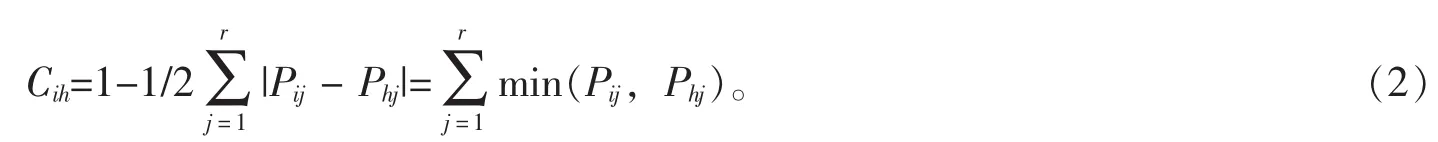
式(2)中: Cih表示物种 i与 h的相似程度, 且有 Cih=Chi, 式(2)具域值[0,1];Pij, Phj分别为物种 i和物种h在资源位j上的重要值百分率。
生态位重叠是指一定资源序列上,2个物种利用同等级资源而相互重叠的情况,其测算公式[13-14]:

式(3)(4)中:Lih为物种i重叠物种h的生态位重叠指数;Lhi为物种h重叠物种i的生态位重叠指数;B(sw)i和 Lhi=B(sw)h具有域值[1/r, 1]; Lih, Lhi具有域值[0, 1]。
3 结果与分析
3.1 物种重要值
对喀斯特区灌草群落演替阶段主要种群葛藤的群落进行调查研究,在9个样地中出现植物种共42种。根据重要值计算,其中出现频率较高的是葛藤Pueraria lobata,香叶树Lindera communis,火棘Pyracantha fortuneana,竹叶椒Zanthoxylum planispinum,异叶鼠李Rhamnus heterophylla,粉枝莓Rubus biflorus,其频率均大于44.4%,重要值具体见表1。群落组成类型主要为 “葛藤+香叶树+火棘+异叶鼠李”,其伴生树种除了有薄叶鼠李Rhamnus leptophylla,胡颓子Elaeagnus pungens,小构Broussonetia kazinoki等外,还包括菝葜Smilax china,飞龙掌血Toddalia asiatica,南蛇藤Celastrus orbiculatus,马棘Indigofera pseudotinctoria等多个种。研究发现:在喀斯特区早期群落演替阶段,主要以藤刺灌丛为主,并有多个群落组成类型,而 “葛藤+香叶树+火棘+异叶鼠李”群落类型只是其中1种,也是喀斯特区典型代表。

表1 喀斯特区葛藤种群灌木层主要种重要值Table1 Important value of main species of Pueraria lobata shrub layer in karst area
3.2 生态位宽度
生态位宽度主要反应物种对资源利用的程度,树种生态位宽度越大,其生态适应能力赿强,生存机会越大,分布越广。从表2中可知:在灌草阶段,葛藤群落组成类型中,生态位宽度排在前6位的分别是葛藤、香叶树、竹叶椒、异叶鼠李、粉枝莓、火棘,在该群落组成类型中,这6个种生态位宽度较宽,说明在喀斯特区特别是在石漠化地区早期恢复阶段,分布较广、数量较多、利用资源充分,成为该群落类型组的优势种。在喀斯特石漠化区灌草群落阶段有很多群落组成类型,但经过多年的研究发现,这些种都是这阶段的常见种、广泛种、优势种。葛藤冠幅较大,特别是在喀斯特石漠化区,岩石裸露率高,葛藤具有生长速度快,覆盖面积大,在早期恢复阶段治理石漠化有较好的效果。因为在恢复早期,有利于遮阳,使一些相对耐荫植物的侵入,与其葛藤共享资源,使得当前整个葛藤群落具有多样性、复杂性和明显的优势种群,各个种群的生态宽度值表明了它们在群落的地位与作用。

表2 喀斯特区葛藤种群灌木层主要树种的生态位宽度值Table2 Ecological niche breadth of main species in Pueraria lobata shrub layer in karst area
3.3 生态位相似性比例
从表3可知:各种对的生态位相似比例Cih在0~1之间不等,说明在 “葛藤+香叶树+火棘+异叶鼠李”群落组成类型中物种利用资源的相似程度差异较大。其可能的原因是在该群落组成类型中,优势种较为明显,而其他伴生种分布无规律,大多呈集群分布,其物种利用资源的程度有较大差异;另一方面是喀斯特石漠化区物种丰富,在同一区域相同物种分布呈随机分布;再一个方面是,在早期阶段,大多物种为阳性树种,由于葛藤覆盖面积较广,一些喜阳的物种无法与之共同生长。表明在灌草阶段,该群落组成类型不稳定,随着演替的进行,优势种的变化较大。对于相似度较大的物种对,利用资源的相似程度较大,如粉枝莓与竹叶椒,相似度为0.9406。
3.4 生态位重叠值
当2个物种利用同一资源或共同占有某一资源要素(食物、营养成分、空间等)时,会出现生态位重叠现象。生态位重叠较大的物种,或具有相近的生态特性,或对环境因子具有互补性要求,即生态位重叠是2个物种在两者与生态因子联系上的相似性[15]。
16个优势种构成120个物种对,对于Lih的生态位重叠值种对有82对(表4),占68.33%,其中主要种群生态位重叠值集中为0~0.05,即0<Lih≤0.01有 12对,占10.0%,0.01<Lih≤0.02有 15对,占12.5%,0.02<Lih≤0.03有12对,占 10.0%,0.03<Lih≤0.04有 9对,占 7.5%,0.04<Lih≤0.05有 14对,占11.6%。对于 的生态位重叠值种对有82对,占68.33%,其中主要种群生态位重叠值集中在0~0.04之间,即0<Lhi≤0.01有13对,占10.8%,0.01<Lhi≤0.02有17对,占14.2%,0.02<Lhi≤0.03有9对,占7.5%,0.03<Lhi≤0.04有18对,占15%,其他生态位重叠分配格局具体见表4。结果表明:16个种群间并未表现出显著的生态位完全重叠,各种群间利用资源的差异性显著,种群对资源的共享趋势并不明显,显示葛藤群落不稳定。从表3可知,优势种群葛藤与其他树种组成种对的生态位重叠值大多为0~0.0978(Lih),反过来为0~0.0595(Lhi),只与薄叶鼠李未形成生态位重叠,葛藤生态位宽度值最大,则与其他种群的生态位重叠的机会也较大,但具有较大生态位宽度种群之间的生态位重叠值未必高,可
能是物种本身生态学特性不同,对资源段位要求不完全一致,从而导致生态位重叠不高,如火棘。

表3 喀斯特区葛藤种群灌木层主要树种生态位Lih,Lhi及Cih值特征表Table3 Characteristic table of the ecological nicheLih,Lhiand Cihof main specie in Pueraria lobata shrub layer in karst area

表4 喀斯特区葛藤种群灌木层主要树种生态位重叠分配格局Table4 Distribution pattern of ecological niche overlap in Pueraria lobata shrub layer in karst area
4 结论与讨论
在喀斯特区 “葛藤+香叶树+火棘+异叶鼠李”群落组成类型中,生态位宽度排在前6位的分别是葛藤、香叶树、竹叶椒、异叶鼠李、粉枝莓、火棘。树种生态位宽度越大,利用资源的能力越大。本研究结果表明,葛藤在喀斯特石漠化生境中的适应能力较强。
在喀斯特区 “葛藤+香叶树+火棘+异叶鼠李”群落组成类型中,各种对的生态位相似比例Cih为0~1,物种利用资源的相似程度差异较大。在灌草群落演替阶段,该群落组成类型不稳定。
种群间的生态位重叠除表征生态相似性外,还反映竞争关系,这与环境条件变化相关。在该群落组成类型中,群落主要树种生态位重叠基本上是种群的生态位宽度越大,与其他种群的生态位重叠的机会越大,反之则越小。通过对葛藤种群生态位研究发现,具有较大生态位宽度种群之间的生态位重叠值未必高,可能是物种本身生态学特性不同,对资源段位要求不完全一致,从而导致生态位重叠不高。
16个种群间并未表现出显著的生态位完全重叠,各种群间利用资源的差异性显著,表明种群对资源的共享趋势并不明显,显示葛藤群落不稳定。
有资料报道关于葛藤生物入侵的问题,通过对葛藤群落调查及栽培示范园的观测,在贵州喀斯特石漠化生境下,如果不进行人工干预 (尤其是苗期的除草),葛藤会被杂草替代。本研究表明:在气温不高、土层不深厚、雨水保不住的喀斯特石漠化区不存在入侵风险。这正好与野外实际调查相符合,在喀斯特石漠化区的演替高级阶段无葛藤等物种的出现。在石漠化地区发展葛藤做饲料需要加大刈割和施肥量,以提高生物产量。
[1]戈峰.现代生态学[M].北京:科学出版社,2002:266-267.
[2]张忠华,梁士楚,胡刚.桂林岩溶石山阴香群落主要种群生态位研究[J].林业科学研究,2009,22(1):63-68.ZHANG Zhonghua, LIANG Shichu, HU Gang.Niche characteristics of dominant populations in Cinnamomum burmannii community on karst hills of Guilin [J].For Res, 2009, 22 (1): 63-68.
[3]李瑞,张克斌,杨晓晖,等.荒漠化草原人工封育区植物生态位研究——以宁夏盐池为例 [J].水土保持研究,2006: 13 (2): 213-216.LI Rui, ZHANG Kebin, YANG Xiaohui, et al.The studies on vegetable niche of prohibited grassland in desertification region: taking Yanchi County, Ningxia Hui Autonomous as an example [J].Res Soil Water Conserv, 2006: 13(2): 213-216.
[4]张璐,苏志尧,陈北光,等.广东石坑崆森林群落优势种群生态位宽度沿海拔梯度的变化[J].林业科学研究,2007, 20 (5): 598-603.ZHANG Lu, SU Zhiyao, CHEN Beiguang, et al.Variations in nche breadth of dominant plant populations along an altitudinal gradient in Shikengkong of Guangdong Province [J].For Res, 2007, 20 (5): 598-603.
[5]HURCHINSON G E.Homage to Santa Rosalia, or why are there so many kinds of animals[J].Am Nat, 1959, 93: 145-159.
[6]SCHONENER T W.Resource partitioning in ecological communities [J].Science, 1974, 185: 27-39.
[7]陈子萱,田福平,牛俊义,等.玛曲高寒沙化草地植物生态位特征研究[J].草地学报,2007,15(6):525-530.CHEN Zixuan, TIAN Fuping, NIU Junyi, et al.Study on characteristics of plant niche in Maqu Alpine desertified meadow [J].Acta Agrestia Sin, 2007, 15 (6): 525-530.
[8]刘建林,夏明忠,罗强,等.攀西野生葛藤资源的开发利用与可持续发展研究[J].中国林副特产,2004,6(37):52-54.LIU Jianlin, XIA Mingzhong, LUO Qiang, et al.The development and utilization of wild Pueraria lobata resource and study on its sustainable development in Panxi Area [J].For Prod Spec China, 2004, 6 (37): 52-54.
[9]张川黔,顾明.贵州极有开发潜力的饲用植物——葛藤[J].贵州农业科学,1998,26(4):9-12.ZHANC Chuanqian, GU Ming.Kudzu bean a forage crop with higher exploitable potentiality in Guizhou [J].Guizhou Agric Sci, 1998, 26 (4): 9-12.
[10]毛富春,赵伯善.野生植物葛藤的研究利用现状及其开发前景[J].西北林学院学报,1995,10(3):88-92.MAO Fuchun, ZHAO Boshan.Utilization status and development prospect of wild plants of Pueraria lobata [J].J Northwest For Coll, 1995, 10 (3): 88-92.
[11]BROOK B W, GRADY J J, CHAPMAN A P.Predictive accuracy of population viability analysis in conservation biology [J].Nature, 2000, 404: 385-387.
[12]STEWART G B, COLES C F, PULLIN A S.Applying evidence-based practice in conservation management; lessons from the first systematic review and dissemination projects [J].Biol Conserv, 2005, 126 (2): 270-278.
[13]王鹏.天然甜槠林乔木层主要树种生态位研究[J].福建林业科技,2006,33(2):58-62.WANG Peng.The niche of main arbor species of Castanopsis eyrei natural forest[J].J Fujian For Sci Technol, 2006, 33(2): 58-62.
[14]胡正华,于明坚.古田山青冈林优势种群生态位特征[J].生态学杂志,2005,24(10):1159-1162.HU Zhenghua,YU Mingjian.Niche characteristics of dominant populations in Cyclobalanopsis glauca forest in Gutian Mountain [J].Chin J Ecol, 2005, 24 (10): 1159-1162.
[15]吴东丽,张金屯,王春乙,等.野生大豆群落主要种群生态位特征研究[J].草地学报,2009,17(2):166-173.WU Dongli, ZHANG Jintun, WANG Chunyi, et al.Niche characteristic of dominant species in the national protected plant, Glycine soja Sieb.et Zucc.community [J].Acta Agrestia Sin, 2009, 17 (2): 166-173.
