一次大强度运动对大鼠骨骼肌泛素蛋白酶体途径基因表达及蛋白质降解的影响
2012-05-12朱荣马延超王瑞元
朱荣 马延超 王瑞元
1温州医学院(浙江 温州 325035) 2洛阳师范学院 3北京体育大学
骨骼肌适应能力很强,废用可萎缩,运动可肥大。肌肉的这种适应依赖蛋白质合成和分解的平衡。已经证实,泛素蛋白酶体途径(ubiquitin-proteasome pathway,UPP)负责细胞内 80~90%的蛋白质周转,降解胞浆未折叠的蛋白和被calpain、caspase途径解离的肌丝,包括 actin 和 myosin[1],是细胞内蛋白降解的重要环节。该途径由泛素(Ubiquitin,Ub)、负责蛋白泛素化的酶(泛素活化酶、泛素结合酶、泛素连接酶、E4、去泛素化酶)以及26S蛋白酶体组成。蛋白质经一系列蛋白泛素化的酶催化,与多个泛素分子结合,形成泛素化蛋白,从而被26S蛋白酶体识别和降解。涉及肌萎缩的实验发现,骨骼肌特有的Muscle Ring Finger 1(MuRF1)、Muscle atrophy F-box protein(MAFbx)mRNA上调增加途径的活性,促进蛋白分解[2]。 运动中骨骼肌 UPP 活性也升高[3]。 我们前期实验证实,大强度运动后26S蛋白酶体活性增高,促使骨骼肌收缩蛋白降解增强[4]。为了进一步了解泛素蛋白酶体途径其它成分的变化,本实验观察了大鼠1次大强度运动后,腓肠肌Ub、MuRF1、MAFbx基因表达,以及泛素化蛋白和3-甲基组氨酸(3-methylhistidine,3-MH)含量的变化,探讨UPP活性在运动骨骼肌蛋白质降解中发挥的作用,为运动骨骼肌蛋白质代谢研究提供参考。
1 材料与方法
1.1 实验动物
雄性SPF级Sprague-Dawley大鼠36只,8周龄,体重190~220 g,购于北京维通利华实验动物技术有限公司,许可证编号:SCXK(京)2007-0001,动物批号:0118785。本实验经北京体育大学实验动物福利伦理审查委员会同意,严格按照北京体育大学动物实验室[动物使用许可证:SYXK(京)2008-0009]规定执行。屏障环境饲养,温度(23±2)℃,相对湿度(65±5)%,昼夜交替时间为12/12小时。每笼4只,自由饮食与饮水,饲料购于北京维通利华实验动物技术有限公司。本研究主要在中国农业科学院基因实验室、北京体育大学骨骼肌机能实验室、解放军总医院化验室完成。
1.2 运动方式
大鼠购进后,在新环境里适应3天,然后跑台运动适应3天,第1天运动量为10 m/min×10 min,第2天运动量为10 m/min×15 min,第3天运动量为15 m/min×15 min,休息1天。取6只作为安静对照,其余大鼠进行1次1小时、25 m/min、坡度5%、相当于75%VO2max 的跑步运动[5]。
1.3 取材
运动0.5小时即刻、运动1小时即刻、运动后1、2、6小时,分别取6只大鼠称重,用2%戊巴比妥钠(0.25 ml/100g体重)腹腔注射麻醉[6],迅速分离大鼠右后肢肌肉,取等量的红、白腓肠肌,用锡纸包好,投入液氮暂存。取材完毕后将标本转移至-80℃冰箱保存,备用。
1.4 试剂与仪器
1.4.1 主要试剂
提取肌组织mRNA的主要试剂:Trizol(美国initrogen 公司),DNase I (deoxyribonuclease I,DNA酶 I,美国 Promega 公司),Rnase inhibitor(RNA 水解酶抑制剂,美国Promega公司)。反转录试剂盒:ReverTra Ace-α-TM First Strand cDNA Synthesis Kit(Code No.FSK-100,日本TOYOBO 公司)。实时荧光定量PCR试剂盒:SYBR Green Realtime PCR Master Mix(Code:QPK-201,日本 TOYOBO 公司)。 肌组织泛素化蛋白含量检测的主要试剂:Ub(P4D1)鼠单克隆抗体IgG1,HRP羊抗鼠 IgG,β-tubulin鼠单克隆抗体,均购自美国Santa Cruz公司。检测肌组织3-MH含量的主要试剂:高氯酸(北京南尚乐化工厂),荧光胺(美国Sigma公司),3-MH标准品(美国Sigma 公司),MOPS[3-morpholinopropanesulfonic acid,3(N-吗啡啉)丙磺酸,美国Promega公司]。
1.4.2 主要仪器
SLY-RTML六道动物跑台(北京硕林苑科技有限公司),高速电动匀浆器(德国IKA公司),ND-100紫外分光光度计(美国Nano Drop公司),PCR扩增仪(美国Bio-Rad公司),TQTM5实时荧光定量仪(美国Bio-Rad公司),垂直电泳槽(北京六一仪器厂),垂直电转槽(美国Bio-Rad公司),电泳仪(瑞典LKB BROMMA公司),YLN-2000亚力恩凝胶影像分析系统 (北京亚力恩机电技术研究所),色谱柱(ZORBAX SB-C18 4.6×150 mm,5 mm,美国 Agilent公司),Agilent 1100高效液相色谱仪(美国Agilent公司)。
1.5 指标测试方法
1.5.1 基因mRNA检测
Trizol提取骨骼肌总RNA,其中加入RNA水解酶抑制剂抑制RNA降解,加入DNA酶I消除基因组DNA,以保证RNA纯度。取1 μl做1.2%琼脂糖凝胶电泳,检测RNA降解程度(见图1)。取1 μl总RNA,用紫外分光光度计测定RNA纯度和浓度。
图1 腓肠肌总RNA电泳图谱
反转录试剂盒将提取的总RNA转录成cDNA。反应体系为 5×RT buffer 4 μl,dNTP (10 mM)2 μl,Oligo(dT)1 μl,Reverse Tra Ace 1 μl,1 μl RNA 水解酶抑制剂,1 μg 总 RNA,加 DEPC 水至 20 μl。 反应程序为 42℃ 30 min,85℃ 5 min,4℃ 5 min, 反应产物于-20℃保存待用。
引物由北京赛百盛基因技术有限公司合成,其序列见表1。
实时荧光定量PCR反应体系为0.5 μl cDNA、0.5 μl上下游引物、8.5μl dd H2O、10μl SYBR Green Mix,总体积为 20 μl。 反应程序为 94℃ 240 s,(94℃20 s,62℃ 20 s,72℃ 10 s)重复 45 次。 以 3-磷酸甘油醛脱氢酶(GAPDH)为内参,每份样品重复3次,采用相对CT值定量分析(2-△△CT)。计算公式如下:△△CT=△CT处理样品-△CT未处理样品,其中△CT=CT测试指标-CT内参。
1.5.2 骨骼肌泛素化蛋白含量测定[7]
肌纤维蛋白的提取:取液氮研磨好的腓肠肌粉末约 100 mg,加入 0.5 ml A 液(20 mM Tris-Hcl(pH 7.4),5 mM EGTA,1 mM DTT,0.5 mM 碘乙酰胺,10 μg/ml E-64,蛋白酶抑制剂片(1 片/50 ml),现用现配),冰浴高速匀浆 (20000 r/min)10 s,4℃1700 g离心 3 分钟,弃上清。沉淀再加入0.5 ml B液(20 mM Tris-Hcl (pH 7.4),5 mM EGTA,250 mM NaCl,1%SDS,0.5 mM碘乙酰胺,10 μg/ml E-64,蛋白酶抑制剂片(1片/50 ml),现用现配,混匀,4℃ 15000 g离心5 min,取上清液保存于-80℃冰箱。
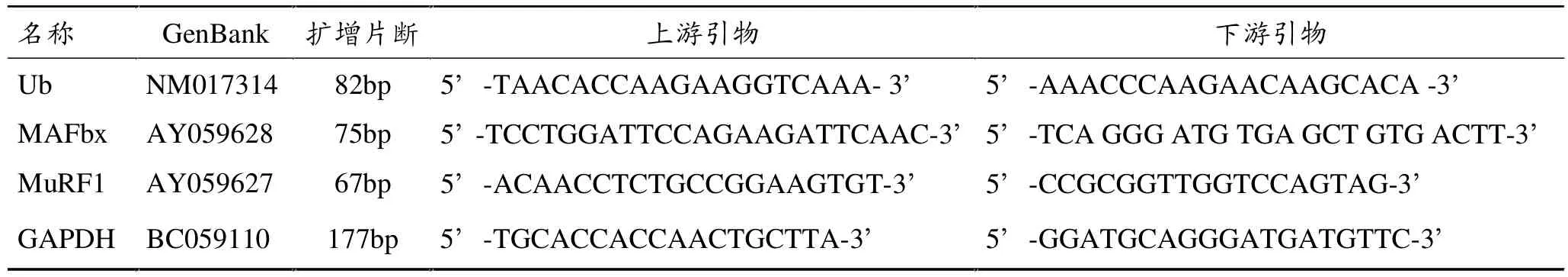
表1 Ub、MAFbx、MuRF-1、GAPDH 引物序列
采用Western Blot方法检测泛素化蛋白含量:蛋白浓度调至2 μg/μl,变性处理后,进行变性聚丙烯酰胺凝胶电泳半定量分析。每孔上样量20 μl,5%浓缩胶90 V电泳1 h,10%分离胶150 V电泳约3.5 h,溴酚蓝泳动到凝胶底部停止电泳。然后300 mA恒流湿转,约1.7 h。3%TBST BSA培养皿中,4℃振摇封闭过夜。 Ub 抗体 1∶1000 稀释,β-actin 抗体 1∶2000 稀释,4℃振摇过夜。 洗膜,再加HRP(1∶2000稀释)室温振摇40分钟。再次洗膜后X线胶片曝光,经影像分析系统拍照,ImageQuant TL软件光密度扫描分析。
1.5.3 骨骼肌 3-MH 含量测定[8,9]
肌肉处理:称取约50 mg腓肠肌,迅速装入离心管(内含预冷3.0%500 μl高氯酸),用剪刀剪碎,冰浴中高速匀浆(20000 r/min)10秒,然后4℃下14000 g离心25 min,提取上清液,备用。
柱前衍生:取50 μl上清液至离心管,加入125 μl 0.2 mM硼酸钠,旋涡振荡,缓慢加入125 μl乙睛(含荧光胺 1.6 g/L),混匀,静置 5 min,加入 18 μl 70%高氯酸,盖紧盖子,80℃水浴1 h。冷却至室温,加入3 M NaOH(含 0.5 M MOPS)50 μl,使标本液 pH 在6.0左右,即可进行高效液相色谱仪检测。
色谱条件:流动相采用10 mM磷酸钠缓冲液(含30%光谱纯乙腈,pH7.5),等度洗脱,流速1.0 ml/min,进样量25 μl,柱温为常温,荧光分光光度计检测,激发 365 nm/发射 460 nm。
色谱分离:3-MH的出峰时间约为5.9分钟,见图2。
标准曲线:3-MH标准品配制成浓度分别为2.5、5.0、7.5、10.0、15.0、20.0 μmol/ml的溶液,与样品处理方法相同,检测对应面积,根据标准品的不同浓度与各自对应的面积建立回归方程,见图3。
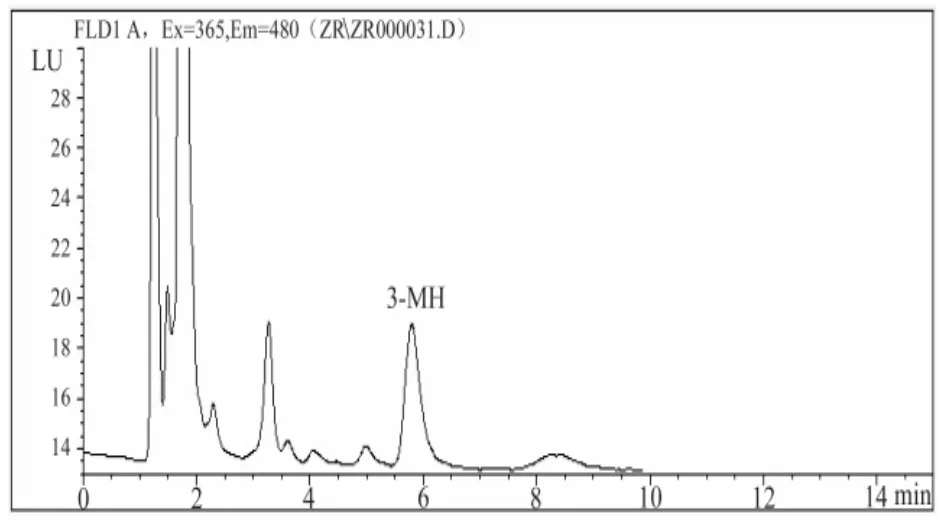
图2 3-MH标准品的高效液相色谱结果
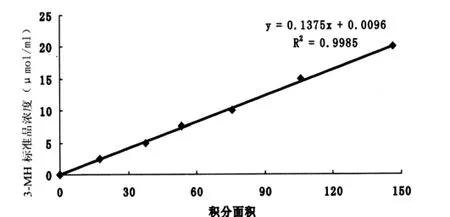
图3 3-MH标准曲线
1.6 统计学分析
用统计学软件SPSS13.0处理数据,进行单因素方差(One-Way ANOVA)分析,结果用平均值±标准差表示,方差齐时用LSD方法,方差不齐时用Tamhane’s方法,P<0.05 为显著性差异,P<0.01为非常显著性差异。
2 结果
2.1 骨骼肌Ub、MuRF1、MAFbx基因表达
表2显示,运动0.5、1小时即刻至运动后6小时,Ub mRNA含量与安静时无显著性差异 (P>0.05)。 1小时运动后 1、2 和 6小 时,MAFbx和MuRF1 mRNA含量较安静时显著升高 (P<0.01,P<0.05),二者运动后2小时表达最高,分别约是安静时的3和4倍,运动后6小时下降,显著低于运动后1小时和 2小时(P<0.05)。
2.2 骨骼肌泛素化蛋白含量
与安静时比较,大强度运动即刻至运动后6小时,骨骼肌泛素化蛋白含量呈逐渐增加趋势(图4、表3),运动后6小时显著高于其它时刻(P<0.01),约是安静时的1.4倍。
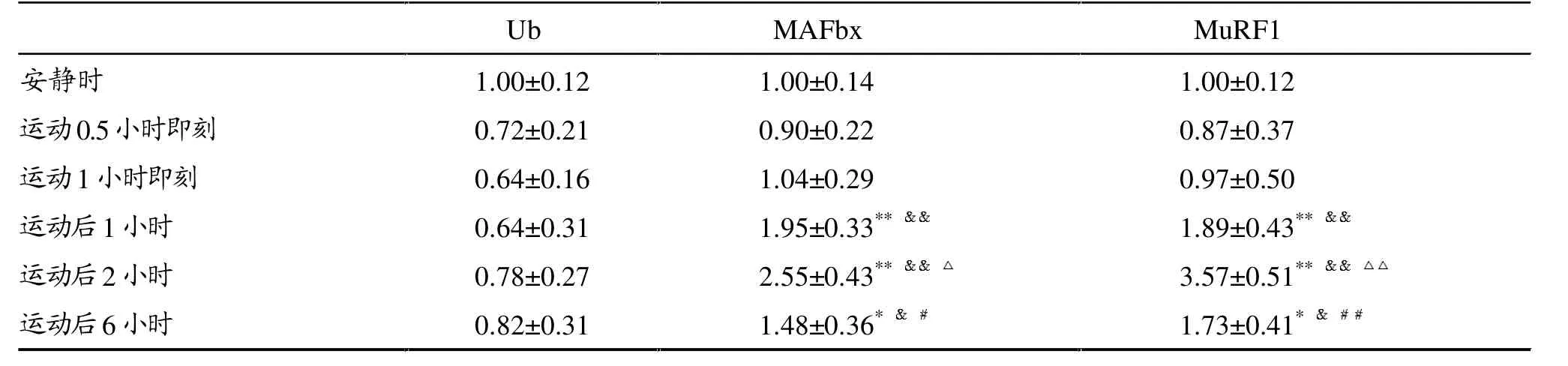
表2 大强度运动后大鼠腓肠肌Ub、MAFbx、MuRF1 mRNA表达变化

图4 腓肠肌泛素化蛋白Western Blot测试图
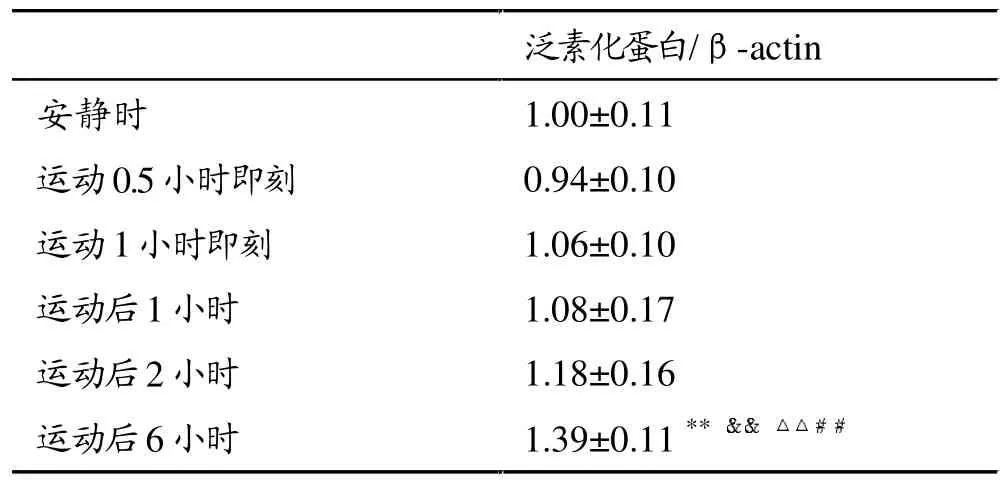
表3 大强度运动后大鼠腓肠肌泛素化蛋白含量变化
2.3 骨骼肌3-MH含量变化
表4显示,运动0.5小时即刻、运动1小时即刻、运动后6小时骨骼肌3-MH含量增加,与安静时比较有显著性差异(P<0.01),其中,运动0.5小时即刻最高,约为安静时的3倍(P<0.01),运动后6小时约为安静时的2倍(P<0.01)。
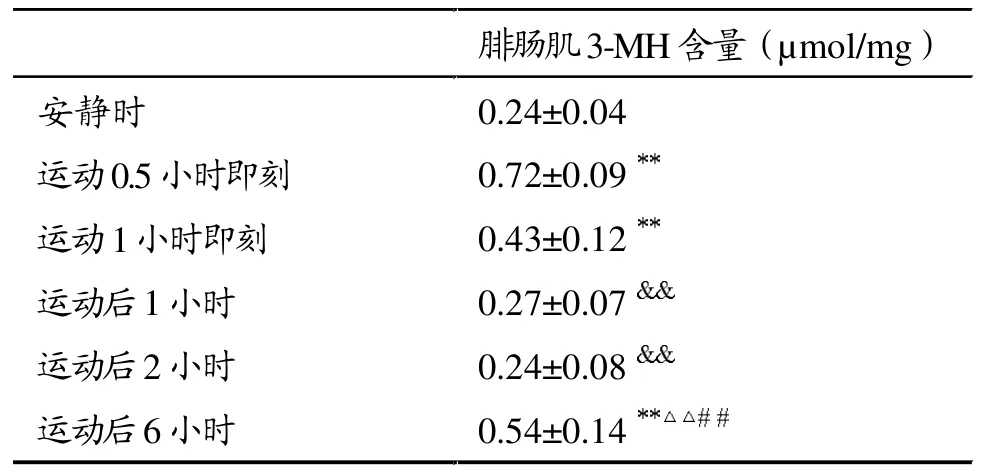
表4 大强度运动后大鼠腓肠肌3-MH含量变化
3 讨论
3.1 大强度运动后腓肠肌泛素蛋白酶体途径基因表达的变化
3.1.1 大强度运动对泛素基因表达的影响
泛素是翻译后修饰蛋白,基因翻译出来的核糖体融合泛素以及多聚泛素,经修饰后才发挥其标记功能。泛素基因的表达被证明与肌萎缩过程同步[10]。Willoughby[11]报道健康志愿者离心伸膝运动后24和48小时,股四头肌泛素蛋白含量和mRNA水平均增加,说明运动加强了泛素基因表达。本研究中大鼠运动骨骼肌泛素mRNA却没有显著性变化,这与Kee等[12]研究结果类似。他认为结果的差异可能与运动形式不同有关;其次,细胞中泛素含量充足,泛素可以循环利用,在短暂的时间内能满足细胞的生理功能。
3.1.2 大强度运动对MuRF1、MAFbx基因表达的影响
MAFbx、MuRF1是骨骼肌里特异性的泛素连接酶E3,催化泛素与底物蛋白结合,在UPP降解骨骼肌蛋白过程中有重要作用。因为在多种减少肌肉使用的模型里,这两个基因表达均显著升高,被认为是肌肉萎缩的显著标志[13]。运动中骨骼肌 MAFbx、MuRF1基因表达也有研究。Mascher等[14]发现不常运动的男青年进行1次膝关节抗阻运动后4~24小时,MuRF1 mRNA含量增加了4.8倍,第2次抗阻运动后MuRF1 mRNA含量比第1次少30%,认为UPP在抗阻运动后肌肉的重塑中起重要作用。Louis等[15]让受试者进行跑步和抗阻运动,采用肌肉活检法发现抗阻运动后1~4小时,腓肠肌和股外侧肌MuRF-1 mRNA含量增加3.5倍,跑步后1~4小时MuRF-1、MAFbx mRNA含量分别增加3.6倍和1.6倍,可见不同形式的运动几乎都能增加MuRF-1、MAFbx基因表达。本研究结果也发现大鼠1小时大强度跑步后 1、2、6 小时, 腓肠肌 MAFbx、MuRF1 mRNA 含量分别是安静时的2倍、2.5倍、1.5倍和1.9倍、3.5倍、1.7倍,与Louis的研究一致。
3.2 大强度运动后腓肠肌3-MH含量的变化
3-MH是组氨酸形成组氨酰-tRNA后发生甲基化的产物,是骨骼肌和心肌蛋白的组成成分[16]。蛋白质代谢中,从多肽链上降解下来的3-MH不能重新合成肽链,也不会被分解,而随尿液排出体外。因此常用尿3-MH含量变化反映骨骼肌蛋白的分解状况。例如,Behm等[17]让受试者做一次最大力量的抗阻运动,运动后3天内肌肉疼痛增加、活动幅度减小,相对疲劳、颤动幅度、尿3-MH含量增多。Bird等[18]发现一组抗阻练习后,受试者尿3-MH含量比练习前有所增加,而其间服用碳水化合物和必需氨基酸时,尿3-MH含量有所减少。但尿3-MH含量并不能真正反映骨骼肌收缩蛋白的降解情况,因为尿中还有非肌组织来源[19]。直接测量肌肉3-MH含量才真正反映骨骼肌收缩蛋白的降解[20]。本实验利用高效液相技术检测运动即刻至运动后6小时的腓肠肌3-MH含量,发现运动0.5小时即刻、运动1小时即刻、运动后6小时骨骼肌3-MH含量较安静时显著增加,说明大强度运动后骨骼肌收缩蛋白有明显降解。张松江[21]利用蛋白组学方法发现大鼠力竭跑后即刻骨骼肌肌球蛋白轻链含量下调,也提示了长时间大强度运动后骨骼肌收缩蛋白降解增强。
3.3 大强度运动后腓肠肌泛素化蛋白含量的变化
泛素化是调节细胞内蛋白质水平的基本方式。泛素经泛素活化酶、泛素结合酶、泛素连接酶以及去泛素酶的依次作用,特异性地标记在蛋白质上,以便被不同细胞器识别,进行一系列生理反应,如细胞周期、细胞凋亡、蛋白转运、抗原提呈和DNA修复,以及细胞对环境刺激的反应等。一般多聚泛素化的蛋白被26S蛋白酶体识别并加以降解。研究表明,运动中肌肉损伤与泛素化蛋白增多有关。如Stupka等[22]发现离心运动后24小时股外侧肌肌力下降,血清CK升高;肌肉活检发现Z盘模糊,泛素化蛋白的含量增加。Dupont-Versteegden 等[23]发现悬吊使大鼠比目鱼肌萎缩,泛素化蛋白含量增加,吊悬加抗阻运动可减小萎缩程度,但泛素化蛋白含量还是增加,这可能与激活了UPP其它成分基因表达,使UPP活性增强有关。本研究也观察到大鼠1小时大强度跑台运动后6小时,骨骼肌泛素化蛋白含量明显增多,这可能也与运动后MAFbx、MuRF1基因表达增加有关,但与肌肉3-MH含量变化的时程不一致,这是因为细胞内存在的泛素化蛋白含量是蛋白泛素化过程和泛素化蛋白降解过程间平衡的反映。当泛素化过程和蛋白降解过程均加速时,不造成泛素化蛋白积累,但降解产物一样增多,只有当泛素化过程快于蛋白质降解时,泛素化蛋白才产生积累。
4 总结
大强度运动后,骨骼肌MuRF1、MAFbx基因表达升高,泛素化蛋白、3-MH含量增加,提示UPP活性增强,并加速骨骼肌收缩蛋白降解。
[1]Ventadour S,Attaix D.Mechanisms of skeletal muscle atrophy.Curr Opin Rheumatol,2006,18(6):631-635.
[2]Altun M,Besche HC,Overkleeft HS,et al.Muscle wasting in aged,sarcopenic rats is associated with enhanced activity ofthe ubiquitin proteasome pathway.JBiolChem,2010,285:39597-39608.
[3]Yang Y,Jemiolo B,Trappe S.Proteolytic mRNA expression in response to acute resistance exercise in human single skeletal muscle fibers.J Appl Physiol,2006,101(5):1442-1450.
[4]朱荣,马延超,许寿生,等.一次大强度运动后大鼠骨骼肌收缩蛋白降解和26S蛋白酶体活性的变化.中国运动医学杂志,2010,29(3):305-308.
[5]Ji LL,Gomez-Cabrera MC,Steinhafel N,et al.Acute exercise activates nuclear factor (NF)-κB signaling pathway in skeletal muscle.FASEB J,2004,18:1499-1506.
[6]孙靖.实验动物学基础.北京:北京科学技术出版社,2005.182.
[7]Vermaelen M,Marini JF,Chopard A,et al.Ubiquitin targeting of rat muscle proteins during short periods of unloading.Acta Physiol Scand,2005,85:33-40.
[8]申传安,柴家科,廖杰,等.高效液相色谱-荧光检测法测定大鼠骨骼肌组织内微量三甲基组氨酸.军医进修学院学报,2003,24(2):120-122.
[9]Kadowaki M,Harada N,Takahashi S,et al.Differential regulation of the degradation of myofibrillar and total proteins in skeletal muscle of rats:effects of streptozotocin-induced diabetes,dietary protein and starvation.J Nutr,1989,119:471-477.
[10]Medina R,Wing SS,Goldberg AL.Increase in levels of polyubiquitin and proteasome mRNA in skeletal muscle during starvation and denervation atrophy.Biochem J,1995,307(Pt3):631-637.
[11]Willoughby DS,Taylor M,Taylor L.Glucocorticoid receptor and ubiquitin expression after repeated eccentric exercise.Med Sci Sports Exerc,2003,35(12):2023-2031.
[12]Kee AJ,Taylor AJ,Carlsson AR,et al.IGF-1 has no effect on postexercise suppression of the ubiquitin-Proteasome system in rat skeletal muscle.J Appl Physiol,2002,92(6):2277-2284.
[13]Glass DJ.Molecular mechanisms modulating muscle mass.Trends Mol Med,2003,9(8):344-350.
[14]Mascher H,Tannerstedt J,Brink-Elfegoun T,et al.Repeated resistance exercise training induces different changes in mRNA expression of MAFbx and MuRF-1 in human skeletal muscle.Am J Physiol Endocrinol Metab,2008,294(1):E43-E51.
[15]Louis E,Raue U,Yang Y,et al.Time course of proteolytic,cytokine,and myostatin gene expression after acute exercise in human skeletalmuscle.J ApplPhysiol,2007,103:1744-1751.
[16]Kumar V,Atherton P,Smith K,et al.Human muscle protein synthesis and breakdown during and after exercise.J Appl Physiol,2009,106:2026-2039.
[17]Behm DG,Baker KM,Kelland R,et al.The effect of muscle damage on strength and fatigue deficits.J Strength Cond Res,2001,15(2):255-263.
[18]Bird SP,Tarpenning KM,Marino FE.Liquid carbohydrate/essential amino acid ingestion during a short-term bout of resistance exercise suppresses myofibrillar protein degradation.Metabolism,2006,55(5):570-577.
[19]Trappe T,Williams R,Carrithers J,et al.Influence of age and resistance exercise on human skeletal muscle proteolysis:a microdialysis approach.J Physiol,2004,554:803-813.
[20]申传安,柴家科,姚咏明,等.胰岛素强化治疗对烫伤脓毒症兔骨骼肌蛋白高降解的调节及其机制.中国危重病急救医学,2006,18(3):139-142.
[21]张松江.急性力竭运动大鼠腓肠肌的蛋白质与蛋白组差异性表达的研究.湖南师范大学硕士学位文,2006.41.
[22]Stupka N,Tarnopolsky MA,Yardley NJ,et al.Cellular adaptation of repeated eccentric exercise-induced muscle damage.J Appl Physiol,2001,91(4):1669-1678.
[23]Dupont-Versteegden EE,Fluckey JD,Knox M,et al.Effect of flywheel-based resistance exercise on processes contributing to muscle atrophy during unloading in adult rats.J Appl Physiol,2006,101:202-212.
