PLIN基因多态性在汉族肥胖儿童青少年中的分布及其与BMI的关系
2012-05-12刘阳陈燕波陈志民常翠青
刘阳 陈燕波 陈志民 常翠青
北京大学第三医院运动医学研究所(北京 100191)
近年来,儿童青少年肥胖患病率迅猛增长,不仅对他们身心健康产生影响,还会对其成年后的健康产生危害[1-3]。研究表明,除不良生活方式外,常见基因突变或基因多态性 (single nucleotide polymorphism,SNP)在肥胖的流行中也发挥着重要作用[4,5]。基因多态性与体重之间关系的研究日益受到重视。
2005年,PLIN基因被纳入为肥胖相关基因。人PLIN基因定位于15号染色体2区6带1亚带(15q26.1)[6],一个与高血压、高甘油三酯血症及糖尿病发生有关的连锁区内[7,8]。PLIN编码的脂滴包被蛋白Perilipin(又称周脂素)是脂滴表面含量最多的一种可磷酸化蛋白,它对脂滴内甘油三酯的储存和动员具有关键性的调节作用[9]。目前发现,PLIN基因在生物体共有79个单核苷酸多态性,其中人类有76个。对PLIN基因多态性与人类肥胖的相关研究发现,PLIN 6209T>C、PLIN 11482G>A、PLIN 13041A>G和PLIN 14995A>T四个位点的SNPs可能与成人肥胖相关,且有种族和性别差异[10-12]。另有研究显示,PLIN 11482G>A与运动、饮食或药物干预后体重的调节有关[13-15]。本研究以北京市200名肥胖中小学生为对象,初步探讨PLIN基因多态性在中国汉族肥胖儿童青少年中的分布及其与BMI的关系。
1 对象与方法
1.1 研究对象
通过门诊招募广告征集和健康体检,筛选出200名10~17岁汉族肥胖儿童青少年,其中男生110名,年龄(14.8±1.8)岁;女生 90 名,年龄(14.7±1.7)岁。肥胖判断依据“中国学龄儿童青少年超重、肥胖筛查体重指数值分类标准”[16]。所有受试者均无吸烟、饮酒史和减肥药物使用史,排除心、肺、肝、肾等重要脏器疾病、身体发育异常、身体残缺、畸形,以及内分泌疾病、药物副作用等引起的继发性肥胖。本研究得到北京大学医学部伦理委员会批准,所有受试者均自愿参加,受试者及其监护人签署知情同意书。
1.2 体格测量
采用立柱式身高坐高仪测定身高。受试者赤脚,立正,躯干自然挺直,头部正直,足跟、骶骨及两肩胛间三点与立柱相接触,测量人员两眼平视水平刻度,测量数据精确到0.1 cm;杠杆秤称量体重,精确到0.1 kg。要求受试者空腹、脱鞋、只穿轻薄衣物。根据体重和身高计算BMI,单位为kg/m2。用MC-180身体成分分析仪 (Tanita公司)测量体脂百分比(FAT%)。
1.3 采集血样标本
清晨采集空腹肘静脉血4 ml,EDTA抗凝,分离血浆和血细胞后,放入-80℃冰箱备用。
1.4 DNA提取和基因组测序
取-80℃冻存EDTA抗凝血细胞样本0.5 ml,使用蛋白酶消化法提取DNA(天根生化科技公司血液基因组DNA提取试剂盒)。
使用Primary Primers 5.0分别设计PLIN 6209T>C(rs2289487)、PLIN 11482G>A(rs894160)和 PLIN 14995A>T(rs1052700)的引物,具体引物序列见表1。聚合酶链反应 (PCR)扩增DNA,PCR反应体系50 μl:其中 2×PCR mastermix × 25μl(天根生化科技公司),上下游引物各 10 μmol/L,模板 DNA1 μg,去离子水加至50 μl。扩增反应在Gene Amp PCR system 9700仪上进行。 反应参数分别为:94℃ 3 min;94℃30 s,69℃ 30 s (6209T>C)、59℃ 30 s (11482G>A)、60℃ 30 s (14995A>T),72℃ 3 min, 共 35 个循环;72℃7 min。PCR产物经常规切胶纯化后由北京擎科新业生物技术有限公司使用ABI3730XL测序仪测序,用Chromas软件分析测序结果。
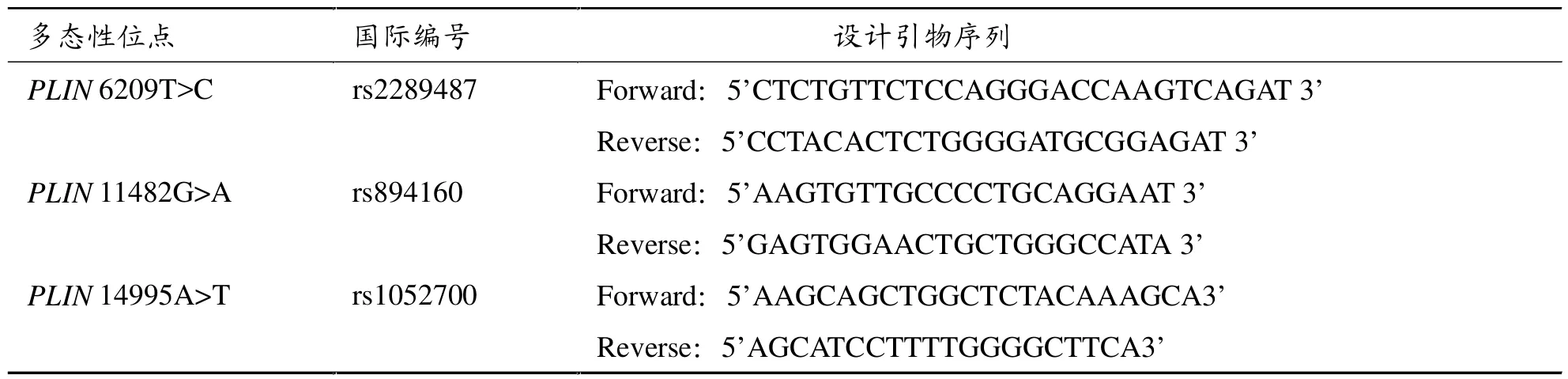
表1 PLIN基因三个多态性位点的引物设计
1.5 统计学分析
所有资料均采用SPSS 17.0统计。符合或近似正态分布的计量资料均以均数±标准差()表示。对基因位点分布情况进行Hardy-Weinberg平衡检验;采用Haploview进行连锁不平衡(Linkage disequilibrium,LD)分析,并计算出连锁不平衡系数D和D’值。为进一步分析PLIN基因多态性与体重及BMI的关系,分析由PLIN 6209T>C、PLIN 11482G>A和PLIN 14995A>T两两组合成的单倍型(haplotype),即两个SNP罕见基因的携带者和非携带者,如6209C/11482A罕见基因CA的携带者 (同时携带PLIN 6209 CC/TC 和 PLIN 11482 GA/AA 的受试者为CA携带者)和非CA携带者。两组之间比较采用独立样本t检验,多组之间的比较采用方差分析(ANOVA)。所有统计检验均采用双侧检验,P<0.05被认为所检验指标有显著性差别。
2 结果
2.1 受试者一般情况
表2显示,受试者平均年龄14.8岁(10~17岁),无性别差异。身高、体重、BMI和体脂百分比的性别差异显著(P<0.05),男生平均身高、体重和BMI均高于女生,FAT%小于女生。
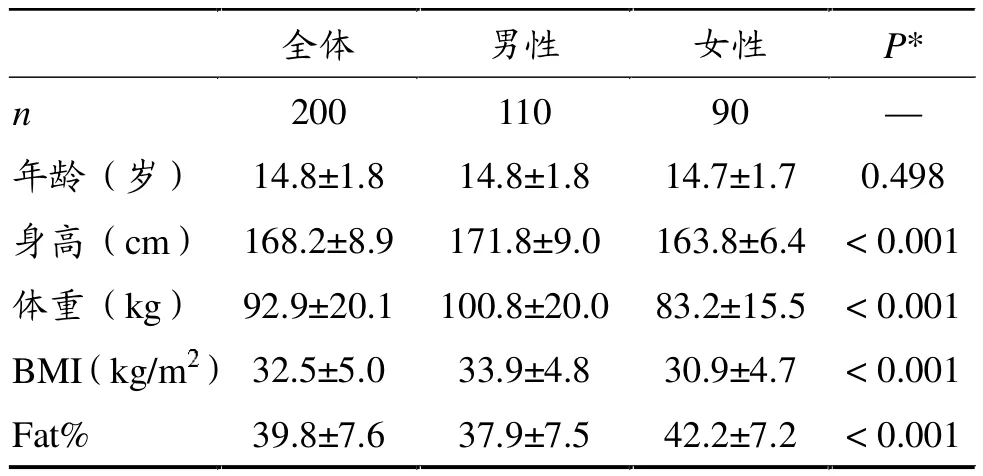
表2 受试者体格测量指标
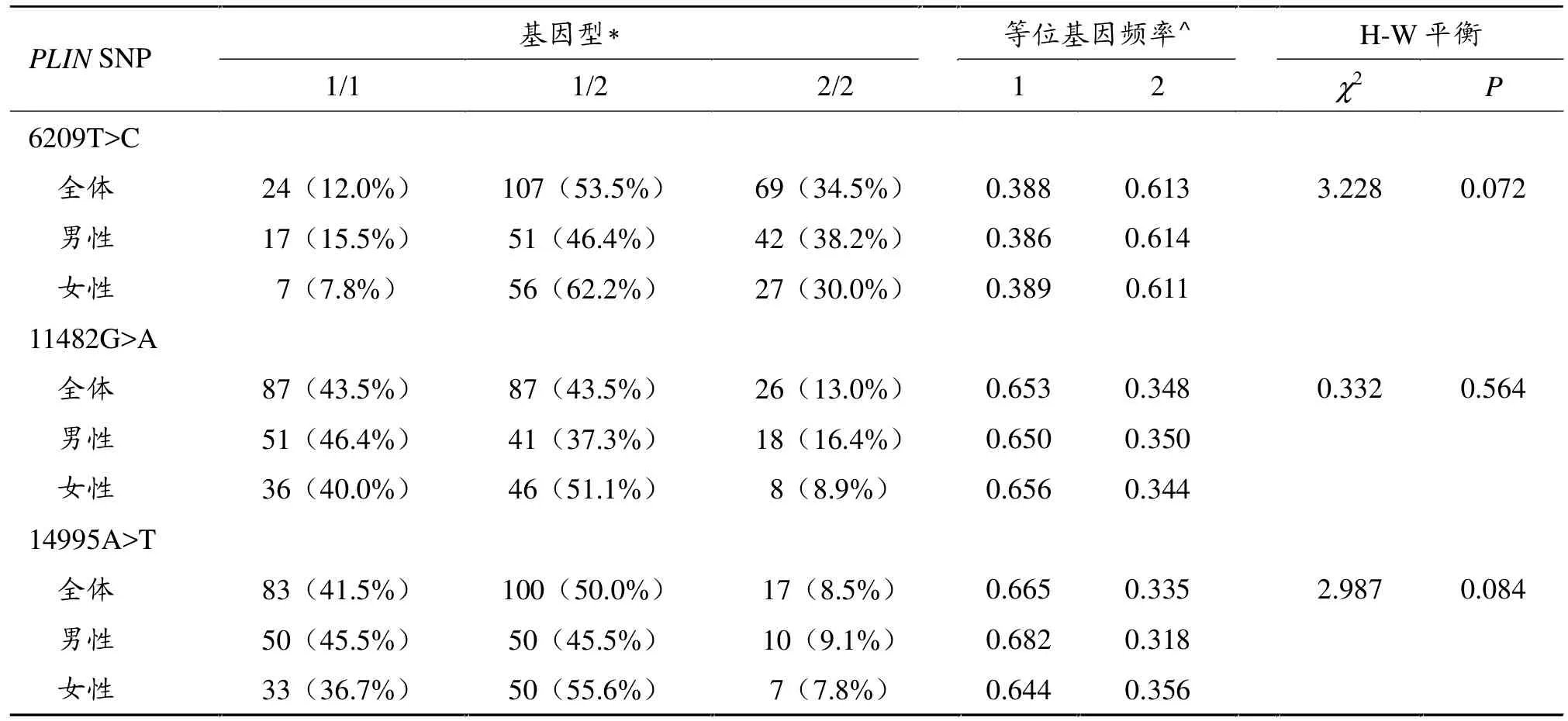
表3 PLIN基因三个多态性位点在男女之间的基因型和频率分布
2.2 PLIN基因多态性位点的频率分布及Hardy-Weinberg平衡检验
表3显示,在所有受试者中,PLIN 6209T>C、PLIN 11482G>A和PLIN 14995A>T的分布情况以及Hardy-Weinberg平衡检验结果显示,群体基因遗传平衡,所选样本均具有整体代表性 (P>0.05)。PLIN 6209T>C、PLIN 11482G>A 和 PLIN 14995A>T多态性罕见基因的频率分别为 0.613、0.348和0.335。卡方检验显示男性和女性之间的PLIN基因多态性的基因型/频率分布无显著差异。
2.3 PLIN基因多态性位点连锁不平衡检验
图1中,左上方块表示PLIN 6209T>C和PLIN 11482G>A之间的LD程度,右上方块显示PLIN 11482G>A和PLIN 14995A>T之间的LD程度,下面的方块表示PLIN 6209T>C和PLIN 14995A>T之间的LD程度。在受试者中,PLIN 6209T>C和PLIN 11482G>A多态性强连锁不平衡或紧密连锁 (D’=0.923)。PLIN 6209T>C和 PLIN 11495A>T及 PLIN 11482G>A和PLIN 11495A>T轻度连锁不平衡,D’值相对较低,分别为0.792和0.727。

图1 PLIN 6209T>C(rs2289487)、PLIN 11482G>A(rs894160) 和 PLIN 14995A>T(rs1052700)3个SNP位点LD图
2.4 PLIN基因多态性与体格测量指标的关系
表4显示,在所有受试者中,PLIN基因3个多态性(包括PLIN 6209T>C、11482G>A和14995A>T)各基因型携带者之间的体重、BMI及Fat%均无显著性差异。按性别分类统计显示,女性受试者中,PLIN基因3个多态性各基因型携带者之间的体格测量指标无显著性差异;而在男性受试者中,PLIN 11482G>A罕见基因携带者(GA+AA)BMI显著高于常见基因(GG)携带者(P<0.05)。

表4 PLIN 11482G>A不同基因型体格测量指标之间的比较
2.5 单倍型分析
表5显示,在所有受试者中,2个SNP组合成的单倍型之间的体格测量指标无统计学差异。按性别分类统计后,在单倍型PLIN 6209T>C/PLIN 11482G>A中,男性受试者中,罕见基因即CA单倍型携带者 (同时携带PLIN 6209 CC/TC和PLIN 11482 GA/AA的受试者)BMI显著高于非CA携带者(P<0.05),其它无显著差异。
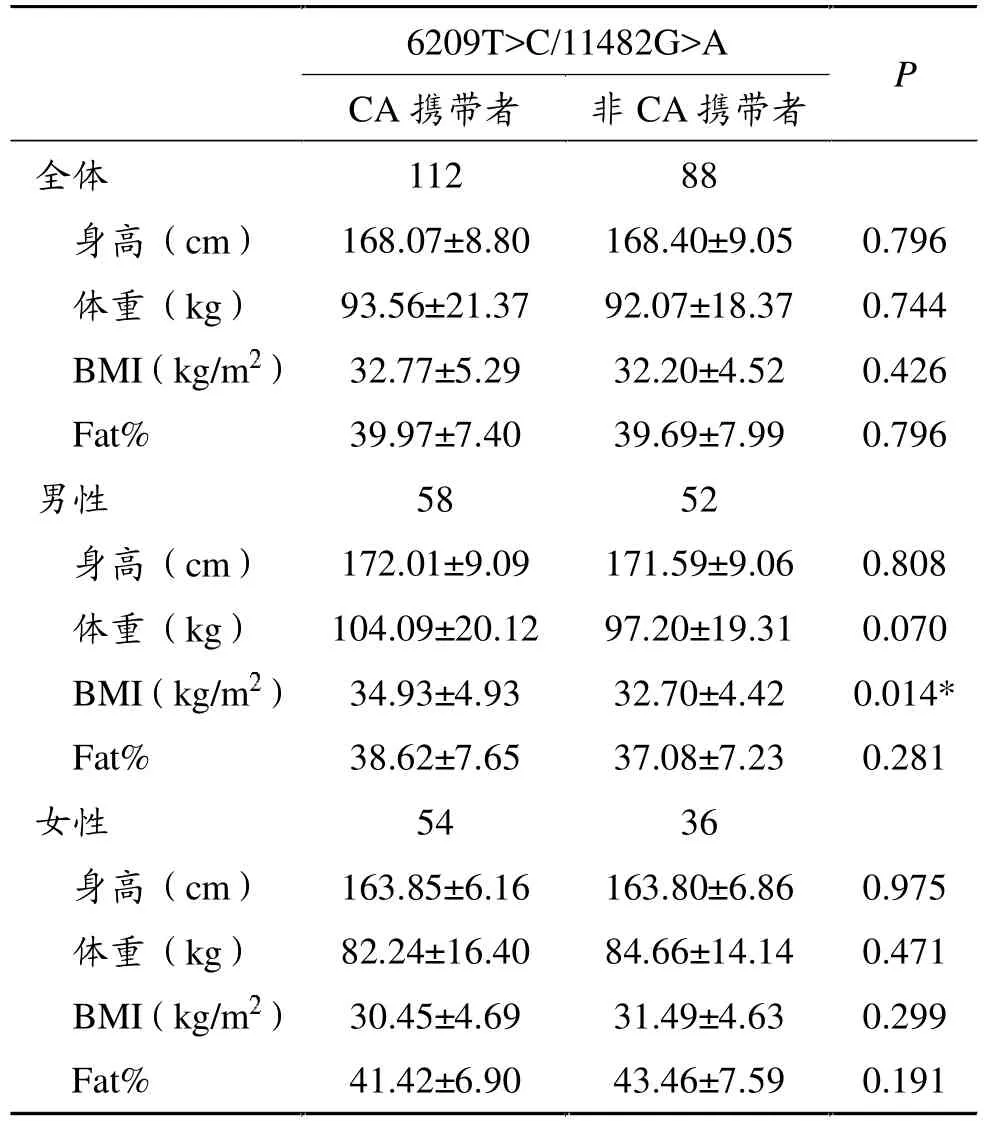
表5 PLIN 6209T>C/PLIN 11482G>A单倍型体格测量指标之间的比较
3 讨论
本研究发现,在中国汉族肥胖儿童与青少年中,PLIN 6209T>C以罕见基因C多见,其频率(0.613)高于巴西肥胖儿童青少年(0.48)[17]和中国汉族肥胖成人(0.556)[18],高于韩国超重或肥胖成人(0.359)[19]和西班牙肥胖成人(0.34)[13];PLIN 11482G>A 以常见基因G为主,其频率(0.653)接近巴西肥胖儿童青少年(0.70)[17],与中国肥胖成人(0.642)[18]及韩国肥胖成人(0.672)[19]相当,低于西班牙肥胖成人(0.76)[13];PLIN 14995A>T以常见基因A多见,其频率(0.665)低于巴西肥胖儿童青少年(0.74)[17],高于中国肥胖成人(0.612)[18]、韩国肥胖成人(0.616)[19]和西班牙肥胖成人(0.62)[13]。上述结果显示,PLIN基因SNPs在肥胖者的分布频率不仅存在种族差异,而且可能存在生理年龄差异,即PLIN基因SNPs在肥胖儿童青少年和肥胖成年人之间的分布频率可能有差异。
对 PLIN 6209T>C、PLIN 11482G>A 和 PLIN 14995A>T三个位点与肥胖受试者体格测量指标之间的关系分析显示,PLIN 11482G>A罕见基因携带者 (GA+AA)BMI显著高于常见基因GG携带者,罕见基因A可能与男性高BMI相关。Deram等[17]对234名巴西肥胖儿童青少年的研究也有类似发现,PLIN 6209T>C、PLIN 11482G>A 和 PLIN 14995A>T三个基因多态性位点与基线体重、腰围及BMI等无明显相关性,但PLIN 11482G>A罕见基因A携带者甘油三酯(TG)水平明显较高,而高密度脂蛋白胆固醇(HDL-C)明显较低,且罕见基因A还与胰岛素抵抗相关。这说明PLIN 11482G>A罕见基因A可能是肥胖儿童青少年及其相关代谢疾病的危险因素。一项有关日本中年男性的病例对照研究结果显示,PLIN 11482G>A罕见基因A与高体重、高BMI及高腰围相关,AA基因型携带者体重、BMI及腰围均明显高于其他基因型[20]。但在中国汉族肥胖成人[18]与西班牙肥胖成人[13]和普通人群[12]中,PLIN 11482罕见基因A可能是肥胖的保护因素,罕见基因A携带者 (GA+AA)BMI和腰围均明显较低[12,13],特别是女性[12]。上述结果不一致除了种族差异外,可能与生活方式等环境因素有关[21,22]。有研究发现,碳水化合物摄入量较高(>144 g/d)的受试者,PLIN 11482罕见基因 A显示是肥胖的保护因素,而摄入量较低者(<144 g/d),罕见基因A则增加肥胖发生的风险[23]。
为了对PLIN基因多态性与体重及BMI的关系进行进一步的关联分析,根据连锁不平衡的情况,将不同的多态性位点组合成单倍型进行分析,结果发现同时携带PLIN 6209T>C和PLIN 11482G>A(2个SNP强连锁,D’=0.923)罕见基因的男性受试者,即CA单倍型携带者BMI显著高于非CA携带者。研究巴西肥胖儿童青少年中PLIN 11482G>A和PLIN 14995A>T组成的单倍型后发现,携带PLIN 11482罕见基因A的单倍型与肥胖相关的高血脂及代谢综合征高风险有关[17]。Qi等[10]研究了新加坡多民族的普通人群后发现,马来裔和印裔中,携带PLIN 11482罕见基因A和PLIN 14995罕见基因T的单倍型(2个 SNP 强连锁:马来裔,D’=0.78;印度裔,D’=0.76)与肥胖高风险相关。但在荷兰超重或肥胖人群及西班牙、美国普通人群中,携带PLIN 6209罕见基因C和PLIN 11482罕见基因A单倍型 (2个SNP 强连锁:荷兰,D’>0.9;西班牙,D’=0.958;美国,D’=0.92)的受试者体重、BMI、腰围、体脂肪和肥胖发生风险均明显较低,且有明显的性别差异[11,12,24]。这说明PLIN基因多态性与肥胖的风险关系还可能与不同人群连锁不平衡的不同结构有关。
本研究的局限性:虽然Hardy-Weinberg平衡检验结果显示群体基因遗传平衡,所选总体样本均具有整体代表性,但缺少正常体重儿童青少年的结果,因此有待增加正常儿童青少年PLIN基因检测结果进一步验证。此外,PLIN基因多态性与肥胖相关代谢指标之间的关系也有待进一步研究。
综上所述,PLIN基因多态性可能与中国汉族肥胖儿童青少年BMI相关,男性儿童青少年中,PLIN 11482罕见等位基因A携带者发生肥胖的风险可能高于GG基因型携带者。
[1]Ebbeling CB,Pawlak DB,Ludwig DS.Childhood obesity:public-health crisis,common sense cure.Lancet,2002,360(9331):473-482.
[2]Kiess W,Reich A,Muller G,et al.Clinical aspects of obesity in childhood and adolescence--diagnosis,treatment and prevention.Int J Obes Relat Metab Disord,2001,25(Suppl 1):S75-S79.
[3]Li YP,Ma GS,Schouten EG,et al.Report on childhood obesity in China (5) body weight,body dissatisfaction,and depression symptoms of Chinese children aged 9-10 years.Biomed Environ Sci,2007,20(1):11-18.
[4]Ramachandrappa S,Farooqi IS.Genetic approaches to understanding human obesity.J Clin Invest,2011,121(6):2080-2086.
[5]Nguyen DM,El-Serag HB.The epidemiology of obesity.Gastroenterol Clin North Am,2010,39(1):1-7.
[6]Nishiu J,Tanaka T,Nakamura Y.Isolation and chromosomal mapping of the human homolog of perilipin (PLIN),a rat adipose tissue-specific gene,by differential display method.Genomics,1998,48(2):254-257.
[7]Mori Y,Otabe S,Dina C,et al.Genome-wide search for type 2 diabetes in Japanese affected sib-pairs confirms susceptibility genes on 3q,15q,and 20q and identifies two new candidate Loci on 7p and 11p.Diabetes,2002,51(4):1247-1255.
[8]Duggirala R,Blangero J,Almasy L,et al.A major susceptibility locus influencing plasma triglyceride concentrations is located on chromosome 15q in Mexican Americans.Am J Hum Genet,2000,66(4):1237-1245.
[9]Tai ES,Ordovas JM.The role of perilipin in human obesity and insulin resistance.Curr Opin Lipidol,2007,18 (2):152-156.
[10]Qi L,Tai ES,Tan CE,et al.Intragenic linkage disequilibrium structure of the human perilipin gene(PLIN)and haplotype association with increased obesity risk in a multiethnic Asian population.J Mol Med,2005,83(6):448-456.
[11]Qi L,Shen H,Larson I,et al.Gender-specific association of a perilipin gene haplotype with obesity risk in a white population.Obes Res,2004,12(11):1758-1765.
[12]Qi L,Corella D,Sorli JV,et al.Genetic variation at the perilipin (PLIN)locus is associated with obesity-related phenotypes in White women.Clin Genet,2004,66 (4):299-310.
[13]Corella D,Qi L,Sorli JV,et al.Obese subjects carrying the 11482G>A polymorphism attheperilipin locusare resistant to weight loss after dietary energy restriction.J Clin Endocrinol Metab,2005,90(9):5121-5126.
[14]KangES,ChaBS,Kim HJ,etal.The11482G >A polymorphism in the perilipin gene is associated with weightgain with rosiglitazone treatmentin type 2 diabetes.Diabetes Care,2006,29(6):1320-1324.
[15]姜雪,黄志卓,常翠青.Plin基因11482 G/A单核苷酸多态性与汉族肥胖青少年有氧锻炼前后BMI变化关系.中国运动医学杂志,2008,28(6):719-722.
[16]中国肥胖问题工作组,季成叶.中国学龄儿童青少年超重、肥胖筛查体重指数值分类标准.中华流行病学杂志,2004(2):97-102.
[17]Deram S,Nicolau CY,Perez-Martinez P,et al.Effects of perilipin (PLIN) gene variation on metabolic syndrome risk and weight loss in obese children and adolescents.J Clin Endocrinol Metab,2008,93(12):4933-4940.
[18]陈燕波,常翠青,黄志卓,等.PLIN基因多态性在中国汉族成年肥胖者中的分布.营养学报,2011,(1):29-33.
[19]Jang Y,Kim OY,Lee JH,et al.Genetic variation at the perilipin locus is associated with changes in serum free fatty acids and abdominal fat following mild weight loss.Int J Obes(Lond),2006,30(11):1601-1608.
[20]Sone Y,Yamaguchi K,Fujiwara A,et al.Association of lifestyle factors,polymorphisms in adiponectin,perilipin and hormone sensitive lipase,and clinical markers in Japanese males.J Nutr Sci Vitaminol (Tokyo),2010,56(2):123-131.
[21]Ordovas JM.Gender,a significant factor in the cross talk between genes,environment,and health.Gend Med,2007,4(Suppl B):S111-S122.
[22]Korkeila M,Kaprio J,Rissanen A,et al.Effects of gender and age on the heritability of body mass index.Int J Obes,1991,15(10):647-654.
[23]Smith CE,Tucker KL,Yiannakouris N,et al.Perilipin polymorphism interacts with dietary carbohydrates to modulate anthropometric traits in hispanics of Caribbean origin.J Nutr,2008,138(10):1852-1858.
[24]Soenen S,Mariman EC,Vogels N,et al.Relationship between perilipin gene polymorphisms and body weight and body composition during weight loss and weight maintenance.Physiol Behav,2009,96(4-5):723-728.
