不同力竭运动后大鼠心脏传导系统PPARα mRNA和蛋白表达的变化及其在运动性心律失常发生中的作用
2012-05-12常芸杨红霞彭泽胄
常芸 杨红霞 彭泽胄
国家体育总局体育科学研究所(北京 100061)
运动性心律失常越来越引起运动医学领域关注,相当一部分运动性心律失常影响到运动员的身体健康、系统训练以及比赛成绩[1-2]。近年,国内外研究报道运动员心律失常发生率高于正常人,且专项训练年限长的运动员中更常见,运动员退赛退役屡见不鲜[3]。而运动性心律失常的发生机制又极其复杂,涉及众多因素。尽管运动医学界对此进行了广泛的临床观察与调研,迄今,运动性心律失常的发生机制仍未完全阐明。心脏传导系统作为心电活动的控制中心和冲动传导的重要部位,其特殊的组织结构和细胞类型决定其具有不同于普通心肌的心电起搏和传导功能,与各种类型心律失常的发生和发展具有密切关系[4]。
心脏终生不停的搏动依赖于心脏强大的能量代谢系统的结构与功能,通常正常心肌能量的70%来源于脂肪酸氧化,因此,线粒体脂肪酸β氧化对维持心肌能量代谢和泵功能有重要意义。过氧化体增殖物激活型受体α(PPARα)作为心肌脂质和能量代谢的重要调控转录因子,调控着出生后心肌线粒体编码脂肪酸β氧化酶类的基因表达,对维持心肌能量代谢和泵功能有重要作用,而运动对心脏传导系统PPARα的影响鲜有报道。为此,本研究进行了力竭运动对心脏传导系统代谢调控PPARα表达的研究,为阐明运动性心律失常的病理改变与发生机制提供实验依据。
1 材料与方法
1.1 实验对象
健康雄性成年SD大鼠100只,8周龄,体重(220±8)g。国家标准啮齿动物饲料喂养,自由饮食。饲养环境为室温(20±2)℃,光照时间 12小时,相对湿度40%~55%。
1.2 运动负荷
将100只实验大鼠随机分为10组,每组10只。其中一次力竭和2周反复力竭游泳运动各4组:运动后即刻组、4小时组、12小时组和24小时组,相应的安静对照组2组。安静对照组不运动,力竭运动各组大鼠尾部负重为体重的3%,2周反复运动组每周运动6天,每天1次。力竭标准参照Thomas的报道[5]。
1.3 心电图描记
所有大鼠于最后一次游泳训练前称重。力竭运动离开水面后,迅速用热吹风机和干布将其皮毛擦干,仰卧固定,采用ADInstruments LabChart 7记录并分析大鼠综合导联心电图。具体计算指标与方法见先前报道[4]。
1.4 取材
分别于最后一次力竭运动后即刻、4小时、12小时及24小时等不同时相取材,迅速取出心脏,用OCT包埋,液氮骤冷,作全心连续冰冻切片,光镜定位心脏传导系统,运用激光显微切割仪切割,分别收集窦房结细胞或细胞团。
1.5 实时荧光定量PCR检测
采用Trizol法提取总RNA,并逆转录cDNA,存于-20℃备用。通过互联网搜索Genbank查找目标因子结蛋白和内参照β-actin的引物基因序列,应用Primer 5软件进行引物设计(表1),设计的引物由上海生工生物技术有限公司合成。引物扩增目标基因片断长度均小于150 bp,其PCR产物用2%琼脂糖凝胶电泳检测验证引物可用后,再进行实时荧光定量PCR,其中预变性 95℃ 30 s,PCR 反应 95℃ 10 s,60℃30 s,40个循环。检测CT值。利用SDS2.2软件对实时定量PCR数据进行分析处理,并导出文件及图像。利用管家基因对目的基因的表达进行校正,得到相对定量结果(相对数值)。
1.6 免疫荧光检测
采用免疫荧光组织化学方法染色心脏传导系统冰冻切片,采用Leica AD MDW活细胞多维图成像工作站和Leica Qwin图像分析系统对目标因子蛋白荧光强度进行定量,荧光强度用积分灰度表示(IOD),参考阴性对照标本中的荧光强度,灰度值在40~130之间为蛋白阳性表达。

表1 PCR引物序列
1.7 统计学分析
所有数据均用SPSS11.5统计软件进行处理,结果用平均数±标准差表示,组间比较采用多因素方差分析,显著性水平为P<0.05。
2 结果
2.1 心脏传导系统形态观察和心电图变化
两种力竭运动后,均有部分大鼠出现心律失常,心脏肉眼观察有不同程度的充血,且反复力竭游泳运动后,各时相组心脏重量指数均显著高于对照组心脏重量指数(P<0.05),具体结果详见先前报道[4]。
2.2 PPARα mRNA表达
2.2.1 一次力竭运动
如表2所示,一次力竭游泳运动后4小时、12小时心脏窦房结PPARα mRNA表达显著低于对照组(P<0.01,P<0.05)。一次力竭游泳运动后即刻和12小时心脏房室结PPARα mRNA表达显著高于对照组(P<0.01)。浦肯野氏纤维PPARα mRNA表达各时相组间无显著性差异(P>0.05)。
一次力竭游泳运动后心脏传导系统不同部位PPARα mRNA表达存有差异,其中,运动后即刻房室结显著高于窦房结(P<0.05),又显著低于浦肯野氏纤维(P<0.01)。
总体看,一次力竭运动后传导系统窦房结、房室结、浦肯野氏纤维PPARα mRNA表达在运动后4小时下降到低谷。其中,房室结变化较明显。
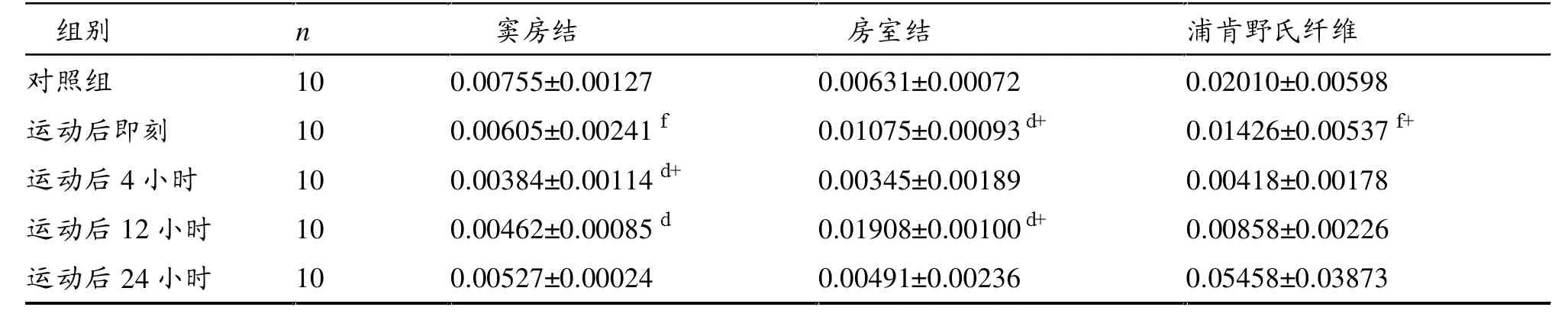
表2 一次力竭运动后大鼠PPARα mRNA相对表达量
2.2.2 反复力竭运动
如表3所示,反复力竭游泳运动后4小时、12小时和24小时心脏窦房结、浦肯野氏纤维PPARα mRNA表达均显著低于对照组和运动后即刻 (P<0.01),其中,运动后即刻组窦房结PPARα mRNA表达又显著高于对照组 (P<0.01),24小时房室结PPARα mRNA表达显著低于对照组(P<0.05)。
反复力竭游泳运动后心脏传导系统不同部位PPARα mRNA表达有差异。运动后即刻房室结PPARα mRNA表达显著高于窦房结 (P<0.05),但低于浦肯野氏纤维(P<0.05),运动后4小时房室结显著高于窦房结和浦肯野氏纤维(P<0.01);运动后12小时房室结显著低于浦肯野氏纤维 (P<0.05),运动后24小时浦肯野氏纤维显著高于窦房结和房室结(P<0.01)。
总体看,反复力竭运动后传导系统窦房结、房室结、浦肯野氏纤维PPARα mRNA表达基本一致。运动后开始下降,4小时到低谷,其中窦房结、房室结变化更明显。
2.2.3 不同力竭方式运动比较
如图1所示,反复力竭后即刻窦房结PPARα mRNA表达显著高于一次力竭后即刻(P<0.05),反复力竭后12小时和24小时又显著低于一次力竭后12 小时(P<0.05)和 24 小时(P<0.01),其他时相组间比较无明显差异(P>0.05)。两种不同方式力竭运动后心脏传导系统房室结PPARα mRNA表达有差异,反复力竭后即刻显著高于一次力竭后即刻(P<0.01),反复力竭后12小时显著低于一次力竭后12小时(P<0.01),其他时相组间无明显差异(P>0.05)。反复力竭后24小时心脏浦肯野氏纤维PPARα mRNA表达显著低于一次力竭后24小时(P<0.01),其他各时相组间无明显差异(P>0.05)。
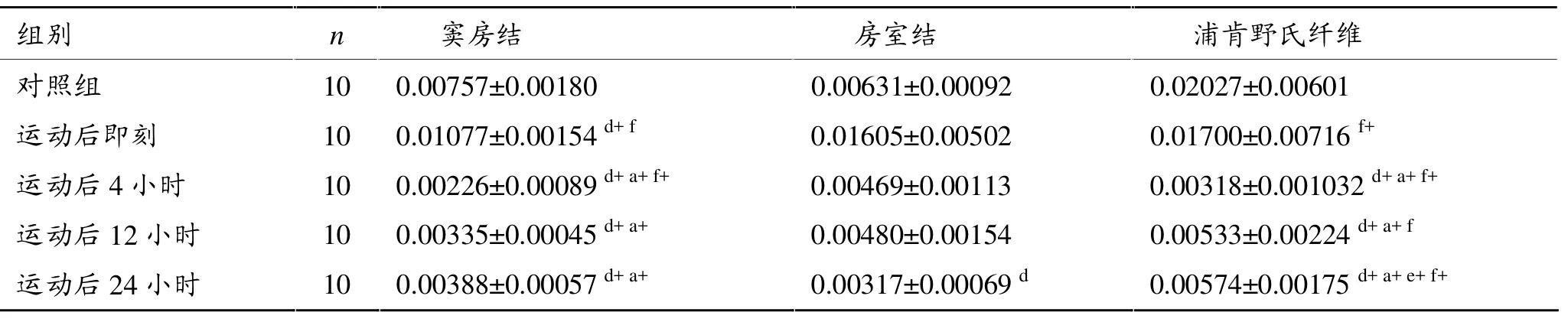
表3 反复力竭运动后大鼠PPARα mRNA相对表达量
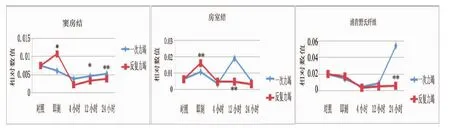
图1 两种不同力竭运动后大鼠心脏传导系统PPARα mRNA相对表达量比较
2.3 PPARα蛋白表达
2.3.1 一次力竭运动
如表4所示,一次力竭游泳运动后12小时心脏窦房结PPARα蛋白表达显著高于对照组和运动后4小时(P<0.05)。运动后即刻、12小时房室结PPARα蛋白表达显著高于对照组 (P<0.01),4小时、24小时显著低于运动后12小时(P<0.01)。运动后即刻、4小时、12小时浦肯野氏纤维PPARα蛋白表达显著低于对照组(P<0.01),4 小时、12 小时显著低于即刻(P<0.01),24小时显著高于对照组和其他各时相组(P<0.01)。
一次力竭游泳运动后心脏传导系统不同部位PPARα蛋白表达存有差异。运动后即刻窦房结PPARα蛋白表达显著低于房室结(P<0.05)和浦肯野氏纤维(P<0.01),房室结显著低于浦肯野氏纤维(P<0.01)。运动后12小时房室结显著高于窦房结和浦肯野氏纤维(P<0.01)。运动后24小时浦肯野氏纤维显著高于窦房结和房室结(P<0.01)。
总体看,一次力竭运动后传导系统窦房结、房室结、浦肯野氏纤维PPARα蛋白表达在运动后4小时下降到低谷。房室结变化较明显,出现两个波峰。运动后浦肯野氏纤维PPARα蛋白表达明显高于窦房结和房室结。
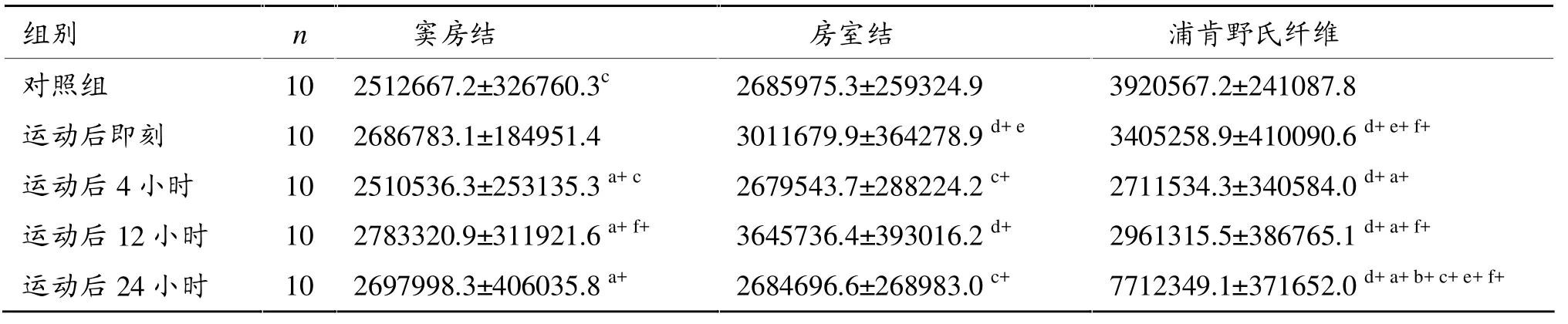
表4 一次力竭运动后大鼠PPARα蛋白表达灰度值变化
2.3.2 反复力竭运动
如表5所示,经反复力竭游泳运动后,心脏传导系统窦房结、房室结PPARα蛋白表达即刻显著高于对照组和其它各运动组(P<0.01)。
浦肯野氏纤维PPARα蛋白表达对照组显著高于其它各运动组(P<0.01),即刻显著高于 4、12、24小时(P<0.01)。
心脏传导系统不同部位在反复力竭游泳运动后存有差异。其中即刻,房室结PPARα蛋白表达显著高于浦肯野氏纤维(P<0.05)。4小时和12小时,窦房结显著低于房室结(P<0.05)和浦肯野氏纤维(P<0.01)。24小时,浦肯野氏纤维显著高于窦房结和房室结(P<0.05)。
表5 反复力竭运动后大鼠PPARα蛋白表达灰度值变化
总体来看,反复力竭运动后传导系统窦房结、房室结、浦肯野氏纤维PPARα蛋白表达规律基本一致。运动后4小时下降到低谷,其中,窦房结、房室结的变化较为明显。
2.3.3 不同力竭方式运动后比较
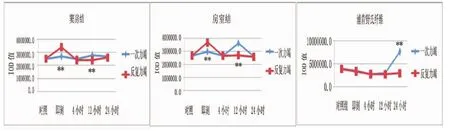
图2 两种不同力竭运动后大鼠心脏传导系统PPARα蛋白表达比较
如图2所示,反复力竭后即刻心脏窦房结PPARα蛋白表达显著高于一次力竭后即刻 (P<0.01),反复力竭后12小时显著低于一次力竭后12小时(P<0.01),其他时相组间无明显差异(P>0.05)。两种不同方式力竭运动后心脏房室结PPARα蛋白表达有差异,反复力竭后即刻显著高于一次力竭后即刻(P<0.01),反复力竭后12小时显著低于一次力竭后12小时(P<0.01),其他时相组间无明显差异(P>0.05)。两种不同方式力竭运动后心脏传导系统浦肯野氏纤维PPARα蛋白表达有差异,反复力竭后24小时显著低于一次力竭后24小时(P<0.01),其他时相组间无明显差异(P>0.05)。
3 讨论
过氧化体增殖物激活型受体(peroxisome proliferator-activated receptor,PPARs)是一种致密分子构成的类固醇受体超家族核受体,有PPARα、PPARβ和 PPARγ3 种亚型[6]。与其它核受体一样,PPARα 在代谢旺盛的组织表达极为丰富,在肝细胞、心肌细胞、肠细胞及肾近曲小管细胞呈高水平表达。其主要功能为介导与脂肪酸氧化有关的基因的活化,使体内脂肪酸氧化/甘油酯化平衡趋向于分解代谢途径;参与调节炎症过程[7]。有研究发现,大鼠心肌缺血缺氧时,脂肪酸氧化减少,葡萄糖氧化增加,减少心肌氧耗,增加心肌对缺血和缺氧的耐受性,阻止细胞凋亡,认为这种能量代谢转换可能具有一定的心肌保护作用[8]。心肌线粒体大部分编码脂肪酸氧化酶核基因的表达是由PPARα调控的,心肌的这种保护作用是通过下调PPARα表达实现的[9]。一旦心肌灌注恢复,PPARα表达未及时恢复,虽然氧供应充足,但脂肪酸氧化未增加,心肌能量供应相对不足,对缺血再灌注的心肌恢复也是不利的[10,11]。 有研究发现,实验性缺血再灌注大鼠心肌PPARα表达减少,同时伴有血清游离脂肪酸增多。还有文献报道,PPARα在心脏还发挥抗炎作用[12],对心脏有保护作用,PPARα激动剂可抑制由脂多糖诱导的肿瘤坏死因子α和核因子κB在心肌的基因表达,提示PPARα激动剂可能通过调控炎症因子发挥抗炎作用,减少心肌受损。
PPARα是心肌脂质和能量代谢的重要调控转录因子,调控出生后心肌线粒体编码脂肪酸β氧化(FAO)酶的大部分核基因表达[13]。研究表明,缺氧时PPARα失活,随之下调FAO酶基因表达。本研究发现,一次力竭运动后窦房结PPARα mRNA与蛋白表达呈先下降后升高的趋势。分析认为,一次力竭运动导致窦房结急性缺氧,PPARα失活,FAO酶基因表达下调,进而使脂肪酸β氧化受到抑制,产能减少。窦房结细胞供能障碍,引起细胞结构和功能的改变,进而引起窦房结起搏和传导功能受限,构成运动性心律失常的发生机制。随力竭运动后恢复时间的延长,PPARα活性逐渐恢复。反复力竭运动后窦房结PPARα mRNA与蛋白表达呈现先升高后下降的趋势,运动后即刻PPARα代偿性升高,有氧氧化代偿性加快,之后随着缺血再灌注加重窦房结损伤,引起PPARα表达相对下降,窦房结细胞严重损伤,且恢复缓慢,至24小时有所回升,且反复力竭后回升速度显著低于一次力竭。这提示反复力竭运动对窦房结的影响较重,可能增加运动性心律失常的发生几率。
本研究发现,一次力竭运动后房室结PPARα mRNA和蛋白表达出现两个波峰。一次力竭运动后即刻PPARα代偿性增加,脂肪酸氧化增强,供能增加。随着缺氧的加重,PPARα活性逐渐降低,曲线下降。氧供恢复后,PPARα大量被激活,有氧氧化能力增强,产能增加。故曲线有所波动。反复力竭运动后呈先升高后下降的趋势。反复力竭运动后呈PPARα代偿性增加,脂肪酸氧化增强,能量代偿性增加,随恢复时间的延长,能量消耗增加,致使缺氧加重,PPARα失活,曲线下降。PPARα表达未及时恢复,虽然氧供应充足,但脂肪酸氧化并未增加,心肌能量供应相对不足。
心肌缺血缺氧时,能量代谢障碍是造成缺血心肌损伤的主要因素[14,15]。 心脏是高氧耗的器官,一旦缺血缺氧,心肌细胞迅速由有氧代谢转为无氧代谢。本研究发现,一次力竭运动后浦肯野氏纤维PPARα mRNA和蛋白表达呈现先下降后升高的趋势,运动后24小时升高最明显。一次力竭运动后PPARα表达失活,可使FAO酶基因表达下调,脂肪酸β氧化受到抑制,产能减少。浦肯野氏纤维细胞供能障碍,可能引起心室内兴奋传导异常。随着运动后恢复时间的延长,PPARα表达被激活,功能逐渐恢复。反复力竭运动后呈下降趋势,运动后4小时明显下降一直持续到24小时,且运动后4小时、12小时、24小时均显著低于对照组和运动后即刻。反复力竭刺激引起PPARα表达持续失活,脂肪酸β氧化持续受到抑制,产能障碍。反复力竭运动后浦肯野氏纤维能量代谢严重不足,其各种功能均受到抑制,室性心律失常发生率大为增加,严重者可能导致心力衰竭。
4 小结
4.1 力竭运动后心脏传导系统各部位能量代谢调控因子PPARα在mRNA和蛋白水平出现异常低表达,易引起传导系统能量代谢障碍,构成运动性心律失常的病理过程和发生机制。
4.2 力竭运动后传导系统各部位PPARα mRNA和蛋白表达有时相规律,不同力竭运动后窦房结、房室结、浦肯野氏纤维PPARα mRNA和蛋白表达均在运动后4小时降至低谷。
4.3 力竭运动后传导系统不同部位PPARα mRNA和蛋白表达存在差异,窦房结PPARα在mRNA和蛋白异常低表达变化规律基本一致,呈持续性低表达,房室结在12小时有所波动,而浦肯野氏纤维在一次力竭运动后24小时PPARα在mRNA和蛋白低表达状况有恢复趋势,进一步提示反复力竭运动对心脏传导系统的影响更为明显。
[1]曲绵域.实用运动医学.北京:北京科学技术出版社,1996.311-318.
[2]常芸.运动心脏理论与实践.北京:人民体育出版社,2008.106-132.
[3]常芸.运动员心脏的医务监督.北京:北京体育大学出版社,2009.203-207.
[4]常芸,杨红霞.不同力竭运动后大鼠心脏传导系统结蛋白mRNA和蛋白表达的变化及其在运动性心律失常发生中的作用.中国运动医学杂志,2012,31(4):314-321.
[5]Thomas DP,Marshall KI.Effect of repeated exhaustive exercise on myocardial subcellular membrane structures.Int J Sports Med,1988,9(4):257-260.
[6]Tyagi S,Gupta P,Saini AS,et al.The peroxisome proliferator-activated receptor:A family of nuclear receptors role in various diseases.J Adv Pharm Technol Res,2011,2 (4):236-240.
[7]Varga T,Czimmerer Z,Nagy L.PPARs are a unique set of fatty acid regulated transcription factors controlling both lipid metabolism and inflammation.Biochim Biophys Acta,2011,1812(8):1007-1022.
[8]Oyekan A.PPARs and their effects on the cardiovascular system.Clin Exp Hypertens,2011,33(5):287-293.
[9]Robinson E,Grieve DJ.Significance of peroxisome proliferator-activated receptors in the cardiovascular system in health and disease.Pharmacol Ther,2009,122 (3):246-263.
[10]Ravingerova T,Adameova A,Carnicka S,et al.The role of PPAR in myocardial response to ischemia in normal and diseased heart.Gen Physiol Biophys,2011,30 (4):329-341.
[11]Hafstad AD,Khalid AM,Hagve M,et al.Cardiac peroxisome proliferator-activated receptor-alpha activation causes increased fatty acid oxidation,reducing efficiency and post-ischaemic functional loss.Cardiovasc Res,2009,83(3):519-526.
[12]Moraes LA,Piqueras L,Bishop-Bailey D.Peroxisome proliferator-activated receptors and inflammation.Pharmacol Ther,2006,110(3):371-385.
[13]PhiliPpe Lefebvre,Giulia Chinetti,Jean-Charles Fruehart,et al,Sorting out the roles of PPARα in energy metabolism and vaseular homeostasis.J Clin Invest,2006,116 (3):571-580.
[14]Rimbaud S,Sanchez H,Garnier A,et al.Stimulus specific changes of energy metabolism in hypertrophied heart.J Mol Cell Cardiol,2009,46(6):952-959.
[15]Madrazo JA,Kelly DP.The PPAR trio:regulators of myocardial energy metabolism in health and disease.J Mol Cell Cardiol,2008,44(6):968-975.
