盐生植物盐地碱蓬质膜Na╋/H╋逆向转运蛋白基因片段的克隆及其序列分析
2012-04-25王生银王锁民
王生银,马 清,王锁民
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020)
盐害是导致农作物减产的主要非生物因素之一[1-2],土壤中高浓度的Na+会扰乱植物正常的代谢活动,影响根系对K+和其他必需元素的吸收,产生渗透胁迫并诱发氧化胁迫等次生胁迫,从而导致植物生长受到抑制,甚至死亡[3-4]。盐生植物在漫长的进化过程中,形成了其特殊的耐盐机制。对其耐盐机理的研究以及耐盐功能基因的挖掘在农作物和优良牧草耐盐性的遗传改良方面有着重要的应用价值。
盐地碱蓬(Suaedasalsa)又名盐蓬、蓬子菜和盐蒿,属藜科碱蓬属一年生多汁草本盐生植物,生于盐渍化土壤、湖滨、河岸和沿海地带,可在400 mmol·L-1的盐浓度下完成其生活史[5],是典型的盐碱地指示植物[6],也是研究植物耐盐机制极好的基因库[7-8]。盐地碱蓬能够在含盐量很高的土壤中正常生长,原因在于其根系吸收的Na+能够有效地运输到植株地上部并储存在液泡中,从而降低植株地上部的水势,提高植株的吸水能力,同时也避免了离子在细胞质中过多积累而扰乱植物正常生理生化代谢活动[9-10]。然而,有关Na+从盐地碱蓬根部向地上部运输机制的研究尚未见报道。近来研究表明,参与Na+从细胞中外排的质膜Na+/H+逆向转运蛋白 (SOS1) 可能参与此过程[11-13]。
质膜Na+/H+逆向转运蛋白的活性首先在大麦(Hordeumvulgare)中发现,并确定其主要与Na+的外排有关[14],是植物抗拒盐离子毒害的首个屏障,质膜 H+-ATPase为其提供能量[15-16]。随后相继从拟南芥(Arabidopsisthaliana)[4,16-18]、水稻(Oryzasativa)[18]、小花碱茅(Puccinelliatenuiflora)[19]等植物中克隆了编码质膜Na+/H+逆向转运蛋白的基因,并确定其参与植物根部Na+的外排[20-22]。然而,Shi等[17]发现,在轻度盐胁迫(25 mmol·L-1NaCl)下,拟南芥质膜Na+/H+逆向转运蛋白 (AtSOS1) 的功能是将Na+装载进入根木质部,以便于Na+向植株地上部转移并最终存贮在叶肉细胞的液泡中;而在重度盐胁迫(100 mmol·L-1NaCl)下,AtSOS1的功能可能是从根的木质部中卸载Na+,从而在盐胁迫下调节Na+通过木质部向植株地上部的运输,减轻过多的 Na+对叶的伤害。Olías等[12-13]研究表明,在盐胁迫下,番茄(Solanumlycopersicum)质膜Na+/H+逆向转运蛋白(SlSOS1)可能通过控制Na+的长距离运输来调控Na+在植物体不同器官中的分布,使Na+大量积累在茎中,从而保护根及重要的光合器官免受Na+毒害。然而,目前对SOS1的研究主要集中在模式植物和甜土植物上,对于盐生植物SOS1的研究相对较少,特别是对积盐型盐生植物的研究尚未见报道。本研究采用 RT-PCR方法克隆积盐型植物盐地碱蓬的SOS1基因片段并分析其序列特征,以期为盐地碱蓬SOS1全长基因的克隆、表达调控等研究奠定基础。
1 材料与方法
1.1试验材料 植物材料为4周龄盐地碱蓬幼苗,种子采自内蒙古自治区查干诺尔湖。大肠杆菌DH5α菌株在草类逆境生理与基因工程实验室保存。
1.2研究方法
1.2.1材料培养 参照马清等[23]的方法。挑选籽粒饱满、无缺损、均匀一致的盐地碱蓬种子,用蒸馏水冲洗2~3遍,在28 ℃下,经蒸馏水浸种催芽24 h。待发芽后,移至灭菌的红沙中,在温室中浇灌Hoagland营养液进行植物材料培养;Hoagland营养液包括6 mmol·L-1KNO3、1 mmol·L-1NH4H2PO4、0.5 mmol·L-1MgSO4·7H2O、0.5 mmol·L-1Ca(NO3)2·4H2O、60 μmol·L-1Fe-citrate、92 μmol·L-1H3BO3、18 μmol·L-1MnCl2·4H2O、1.6 μmol·L-1ZnSO4·7H2O、0.6 μmol·L-1CuSO4·5H2O、0.7 μmol·L-1(NH4)6Mo7)24·4H2O;温室的昼夜温度为(28±2) ℃/(23±2) ℃,光照时间16 h·d-1,光照强度约600 μmol·m-2·s-1,相对湿度60%~80%。
1.2.2总RNA的提取 取150 mmol·L-1NaCl处理48 h的4周龄盐地碱蓬幼苗根系,加入液氮研磨至粉末状,按照UNIQ-10 柱式Trizol总RNA抽提试剂盒的操作说明书提取根系总RNA。用1.0%甲醛变性凝胶电泳鉴定其完整性和质量。
1.2.3引物的设计与合成 参照马清等[24]的方法,通过对其他植物SOS1核苷酸序列进行同源性比较,找出高度保守的区段,根据同源性高和简并性低的原则,利用DNAMAN和Primer 5.0软件设计一对简并引物P1和P2,用于扩增盐地碱蓬SOS1基因片段,推测目的片段的长度为736 bp,引物由上海生工生物工程有限公司合成。
P1:5’- GCATCA(C/T)TT(C/T)TGGGA(A/G)ATGGT -3’;
P2:5’- AT(C/T)CT(C/T/G)CC(C/T)TCATC(A/G)AGCAT -3’。
1.2.4RT-PCR扩增 参照郭强等[24]的方法。PCR扩增反应体系:在200 μL PCR管中依次加入10×PCR Buffer 5 μL、25 mmol·L-1MgCl23.5 μL、2 mmol·L-1dNTP 5 μL、10 μmol·L-1P11 μL、10 μmol·L-1P21 μL、Taq DNA polymerase (5 U·μL-1) 0.5 μL、cDNA 2 μL,加纯水至50 μL。反应条件:94 ℃预变性2 min;94 ℃变性30 s,51 ℃退火50 s,72 ℃延伸60 s,30个循环;最后72 ℃延伸10 min。PCR扩增产物用1.2%琼脂糖凝胶检测,目的片段的回收和纯化按照UNIQ-10柱式DNA 胶回收试剂盒操作说明进行。
1.2.5阳性克隆的筛选与鉴定 参照郭强等[24]的方法。将回收的PCR产物连接到pUCm-T载体,转化感受态大肠杆菌DH5α,用含有50 μg·mL-1氨苄青霉素的LB固体培养基进行蓝白斑筛选,转化白斑菌株经质粒PCR鉴定确认阳性克隆后,送至北京华大基因科技股份有限公司测序。
1.2.6序列分析 序列的比较、翻译等在DNAMAN生物软件上进行,Blast搜索在NCBI (www.ncbi.nlm.nih.gov/BLAST) 网站上进行。
2 结果
2.1RT-PCR扩增 以盐地碱蓬根系为材料提取的总RNA反转录所得到的第一链cDNA为模板,用简并引物P1和P2进行PCR扩增。扩增产物经凝胶电泳检测发现约在736 bp处有1条亮带,且上下无杂带,与目的片段大小一致 (图1),推测可能是盐地碱蓬SOS1基因片段。
2.2阳性克隆的鉴定 将回收纯化的目的片段连接到pUCm-T克隆载体上,转化大肠杆菌DH5α。从转化的平板上随机挑取2个白色菌斑并提取质粒,进行PCR扩增,得到的扩增片段大小约为736 bp(图2),与RT-PCR结果一致,表明这些克隆为阳性克隆。
2.3SOS1基因片段序列分析 测序结果显示,该阳性克隆序列长度为736 bp,推测其编码244个氨基酸 (图3)。Blast比较结果表明,该片段与海松菜(Suaedajaponica)、盐角草(Salicorniabrachiata)、冰叶日中花(Mesembryanthemumcrystallinum)和霸王(Zygophyllumxanthoxylum)等植物SOS1核苷酸序列的同源性均在74%以上,其中与海松菜SjSOS1(AB198179.1)核苷酸序列的同源性高达95%。表明本研究克隆到的片段为SOS1 基因片段,将其命名为SsSOS1。

图1 RT-PCR产物凝胶电泳图
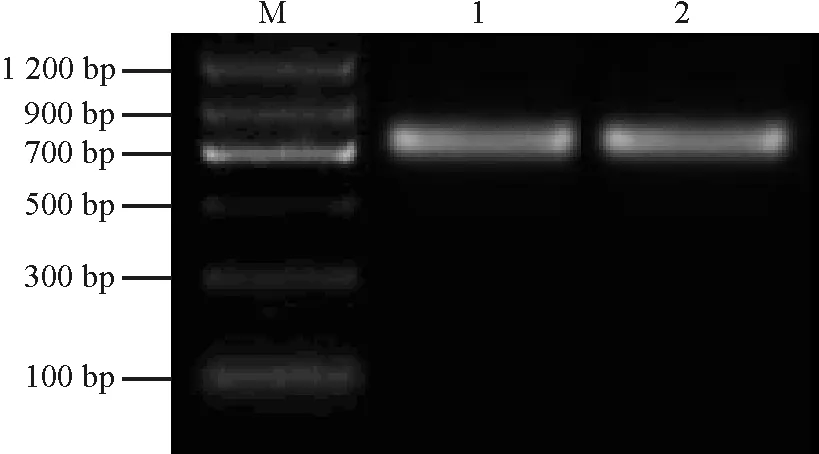
图2 阳性克隆的PCR鉴定

图3 盐地碱蓬SsSOS1核苷酸序列及推测的氨基酸序列
多重比较及系统进化分析表明,盐地碱蓬SsSOS1片段与藜科植物[如海松菜SjSOS1和盐角草SbSOS1(EU879059.1)]的进化关系非常近,氨基酸同源性分别高达98%和90%;与其他双子叶植物如冰叶日中花McSOS1(AM746987.1)和霸王ZxSOS1(GU177864.1)的进化关系相对较远,同源性分别为77%和69%;而与单子叶植物如小花碱茅PtSOS1(GQ452778.1)的进化关系最远,同源性仅为61%(图4、图5)。此外,盐地碱蓬SsSOS1氨基酸序列包含4个跨膜区,相对于非跨膜区而言,跨膜区则具有更高的同源性(图4、图5)。
3 讨论
绝大多数高等植物的质膜Na+/H+逆向转运蛋白均由单基因编码[16,18]。该蛋白N末端为一个疏水结构,主要负责Na+的转运[16],可能由10~12个跨膜结构域组成,不同物种间SOS1的高度同源区域主要位于该端。本研究克隆的SsSOS1片段位于其N末端,包含4个跨膜区,与其他植物SOS1核苷酸的同源性在74% 以上,编码氨基酸的同源性在61%以上,相对于非跨膜区而言,跨膜区具有更高的同源性(图4、图5)。系统进化分析表明,本研究克隆的盐地碱蓬SsSOS1片段与双子叶黎科植物如海松菜SjSOS1和盐角草SbSOS1的进化关系最近,与其他双子叶植物如冰叶日中花McSOS1和霸王ZxSOS1的进化关系相对较远,而与单子叶植物(如小花碱茅PtSOS1)的进化关系最远(图5)。可见,不同植物质膜Na+/H+逆向转运蛋白在进化上具有一定的差异,尤其是在单子叶和双子叶植物之间,这种差异更明显[23]。

图4 SsSOS1氨基酸序列与其他植物SOS1氨基酸序列的多重比较

图5 SsSOS1与其他植物SOS1的系统进化树
SOS1作为植物膜系统的一种转运蛋白,参与细胞内Na+浓度的控制、pH值调节和细胞体积变化等一系列重要的生命活动[25]。除上述功能外,SOS1在控制Na+长距离运输方面起着非常重要的作用。Shi等[17]研究表明,在盐胁迫下,拟南芥质膜Na+/H+逆向转运蛋白(AtSOS1)可以调节Na+在木质部中从根部向地上部的运输,减轻过多的Na+对叶造成的伤害。此外,SOS1在维持植物细胞K+稳态平衡方面起着重要作用,其基因突变和表达下调均会影响植物K+吸收及转运系统的功能,进而破坏细胞K+的稳态平衡[26-27]。同时,SOS1也会影响植物细胞中Ca2+的转运,其活性受抑制后,液泡膜上与Ca2+转运有关的基因表达丰度显著上调,致使液泡中积累大量的Ca2+[28]。可见,SOS1在植物耐盐性方面具有非常重要的作用。本研究成功克隆了SsSOS1基因的核心片段,为SsSOS1全长基因的克隆、表达调控及进一步分析其在盐地碱蓬Na+长距离运输及K+、Ca2+稳态平衡中的作用奠定了基础。
[1]王爱祥,陈爱萍,张博.不同苜蓿品种耐盐性初探[J].草业科学,2010,27(3):102-106.
[2]秦峰梅,张红香,武祎,等.盐胁迫对黄花苜蓿发芽及幼苗生长的影响[J].草业学报,2010,19(4):71-78.
[3]Apse M P,Aharon G S,Snedden W A,etal.Salt tolerance conferred by over expression of a vacuolar Na+/H+antiporter in Arabidopsis[J].Science,1999,285:1256-1258.
[4]Zhu J K.P1ant salt tolerance[J].Trends in Plant Science,2001,6:66-71.
[5]陈敏.盐地碱蓬膜相关蛋白及其在耐盐中的作用[D].济南:山东师范大学,2009:5.
[6]中国科学院《中国植物志》编辑委员会.中国植物志:第二十五卷第二分册[M].北京:科学出版社,1979:115-135.
[7]程国强,王萍.盐地碱蓬耐盐相关基因克隆研究进展[J].生物技术通报,2003(5):18-21.
[8]邹桂梅,苏德荣,黄明勇,等.人工种植盐地碱蓬改良吹填土的试验研究[J].草业科学,2010,27(4):51-56.
[9]Hasegawa P M,Bressan R A,Zhu J K,etal.Plant cellular molecular responses to high salinity[J].Plant Physiology and Plant Molecular Biology,2000,51:463-499.
[10]Song J,Chen M,Feng G,etal.Effect of salinity on growth,ion accumulation and the roles of ions in osmotic adjustment of two populations ofSuaedasalsa[J].Plant and Soil,2009,314:133-141.
[11]Shi H Z,Lee B H,Wu S J,etal.Over expression of a plasma membrane Na+/H+antiporter gene improves salt tolerance inArabidopsisthaliana[J].Nature Biotechnology,2003,21:81-85.
[12]Olías R,Eljakaoui Z,Li J,etal.The plasma membrane Na+/H+antiporterSOS1 is essential for salt tolerance in tomato and affects the partitioning of Na+between plant organs[J].Plant,Cell and Environments,2009,32:904-916.
[13]Olías R,Eljakaoui Z,Pardo J M,etal.The Na+/H+exchangerSOS1 controls extrusion and distribution of Na+in tomato plants under salinity conditions[J].Plant Signaling and Behavior,2009(4):973-976.
[14]Ratner A,Jacoby B.Effect of K+,its coumter anion,and pH on sodium efflux from barley roots[J].Journal of Experimental Botany,1976,148:425-433.
[15]Blumwald E,Aharon G S,Apse M P.Sodium transport in plant cells[J].Biochimica et Biophysica Acta,2000(1465):140-151.
[16]Shi H Z,Ishitani M,Kim C,Zhu J K.TheArabidopsisthalianasalt tolerance geneSOS1 encodes a putative Na+/H+antiporter[J].Proceeding of the National Academy of Sciences of the USA,2000,97:6896-6901.
[17]Shi H Z,Quintero F J,Pardo J M,etal.The putative plasma membrane Na+/H+antiporterSOS1 controls long distance Na+transport in plants[J].The Plant Cel1,2002,14:465-477.
[18]Martínez-Atienza J,Jiang X,Garciadeblas B,etal.Conservation of the salt overly sensitive pathway in rice[J].Plant Physiology,2007,143:1001-1012.
[19]程玉祥.过量表达星星草PtSOS1提高拟南芥的耐盐性[J].植物生理学通讯,2008,44(6):1125-1130.
[20]Oh D H,Leidi E,Zhang Q,etal.Loss of halophytism by interference withSOS1 expression[J].Plant Physiology,2009,151:210-222.
[21]Oh D H,Zahir A,Yun D J,etal.SOS1 and halophytism[J].Plant Signaling and Behavior,2009(4):1081-1083.
[22]Taji T,Seki M,Satou M,etal.Comparative genomics in salt tolerance betweenArabidopsisandArabidopsis-related halophyte salt cress usingArabidopsismicroarray[J].Plant Physiology,2004,135:1697-1709.
[23]马清,包爱科,伍国强,等.质膜Na+/H+逆向转运蛋白与植物耐盐性[J].植物学报,2011,46(2):206-215.
[24]马清,周向睿,伍国强,等.盐生植物碱蓬Actin基因片段的克隆及序列分析[J].生物技术,2009,19(1):1-3.
[25]郭强,周向睿,王沛,等.盐生植物小花碱茅ptAKTL基因片段的克隆序列分析[J].草地学报,2010,18(5):683-688.
[26]Peng Y H,Zhu Y F,Mao Y Q,etal.Alkali grass resists salt stress through high K+and an endodermis barrier to Na+[J].Journal of Experimental Botany,2004,55:939-949.
[27]Qi Z,Spalding E P.Protection of plasma membrane K+transport by the salt overly sensitivel Na+/H+antiporter during salinity stress[J].Plant Physiology,2004,136:2548-2555.
[28]Oh D H,Lee S Y,Bressan R A,etal.Intracellular consequences ofSOS1 deficiency during salt stress[J].Journal of Experimental Botany,2010,61:1205-1213.
