20份马蹄金种质资源遗传多样性RAPD和SRAP分析
2012-04-25周玉锋干友民王小利张建波
付 薇,周玉锋,干友民,王小利,张建波,
(1.贵州省草业研究所,贵州 贵阳 550006;2.四川农业大学草业科学系,四川 雅安 625014)
马蹄金(Dichondrarepens)植株低矮,叶形美观,成坪后无需修剪,管理成本低廉,用途多样,在国外通常用作优良地被绿化材料或固土护坡植物,有很高的推广利用价值[1]。国内许多学者对马蹄金的生物学特征、栽培管理、生理生化等方面开展了相关的研究工作[2-5],但在资源收集、鉴定与评价等方面鲜见报道,分子水平的研究更为少见。我国野生马蹄金群落分布范围较广,受地形、地势及气候差异等多方面因素影响,各地的野生马蹄金在形态及遗传上都存在不同程度变异,是一种极具开发价值的地被植物种质资源四川农业大学干友民等广泛收集四川、云南、贵州、重庆境内的野生马蹄金种质资源,在国内首次对其遗传多样性、形态、生长习性、生产繁殖性能及繁殖技术、抗胁迫能力、景观性能等方面进行评价与筛选,发现野生马蹄金具有优良的绿化价值和生态特性,且各地区的材料在外部形态上和抗性方面存在较大的差异[6-9]。
随着分子标记技术的不断发展成熟,目前已有多种分子标记技术(RAPD、ISSR、SSR和SRAP)被广泛应用于植物遗传多样性分析[10-12]、遗传连锁图谱构建[13-14]、基因定位[15]和比较基因组学研究[16]等方面。利用单一分子标记技术对马蹄金种质遗传多样性进行研究会因为这种分子标记技术本身的局限而使结果不够完整可靠。为此,本研究结合不同分子标记技术的自身特点,以1份美国进口品种为对照,采用RAPD和SRAP 两种分子标记分别对来自四川、重庆、贵州、云南的20份野生马蹄金和1份美国进口品种进行分子水平遗传多样性研究,以期探明马蹄金的遗传基础及不同材料间的遗传差异,获得较为完整的遗传分类及亲缘关系资料,为马蹄金种质资源保护和品种选育提供理论依据。
1 材料与方法
1.1供试材料 20份供试材料来自云南、四川、重庆、贵州等西南地区4省(市),同时以1份美国普通马蹄金(Common)为对照品种。全部资源保育于四川农业大学草业科学系资源圃内和贵州省草业研究所独山基地。
1.2试验方法
1.2.1马蹄金DNA提取 采集新鲜幼嫩的马蹄金叶片,采用广州达晖生物技术公司生产的植物基因组提取试剂盒进行基因组DNA提取。以已知浓度的标准DNA与马蹄金样本DNA在0.8%琼脂糖凝胶上进行电泳,检测DNA质量、浓度,并稀释至10 ng·μL-1,保存于-20 ℃冰箱备用。

表1 试验马蹄金材料及采集地
1.2.2RAPD反应程序 参照张西西等[17]、田志宏等[18]的研究方法,从100个随机引物(由北京赛百盛公司合成)中筛选出20个条带清晰,多态性好的引物对马蹄金DNA进行扩增,扩增体系如下:10×PCR buffer 2.5 μL,模板DNA 30 ng,Mg2+3.5 mmol·L-1,dNTP 300 μmol·L-1,引物0.4 μmol·L-1,TaqDNA聚合酶1.0 U,总体积为20 μL。反应程序:94 ℃预变性5 min;前38个循环程序为94 ℃ 30 s,38 ℃ 30 s,72 ℃ 90 s;最后72 ℃延伸7 min,4 ℃保存。扩增结束后,在1.5%琼脂糖凝胶中电泳,经0.1% 溴化乙锭染色后,凝胶成像系统照相。
1.2.3SRAP反应程序 参照Li和Quiros发表的引物[19]和袁学军等的方法[20],从80对引物(由上海捷瑞生工合成)中筛选出22对进行DNA扩增,优化后扩增体系如下:Mg2+2.5 mmol·L-1、dNTP 0.3 mmol·L-1、引物0.4 μmol·L-1、TaqDNA聚合酶1.0 U,模板DNA浓度10 ng·μL-1,总体积为20 μL。扩增程序:94 ℃预变性5 min;94 ℃变性1 min,35 ℃退火1 min,72 ℃延伸,5个循环;94 ℃变性1 min,52 ℃ 1 min,72 ℃延伸1 min,37个循环;72 ℃延伸10 min,4 ℃保存。扩增结束后,上样于6%的变性聚丙烯酰胺凝胶进行分离,IXTBE缓冲液,电泳结束后快速银染检测。
1.3数据统计与分析 电泳结果数字化及多态性判断,有多态带记为1,无带记为0,将所有材料的扩增条带转换为0,1矩阵后,采用NTSYS-pc软件计算遗传相似系数(GS),GS=2Nij(Ni+Nj),式中,Ni为材料i的条带数,Nj为材料j的条带数,Nij为i和j中都出现的条带数。原始数据的整理采用Excel软件完成,利用NTSYS-pc 2.10软件进行遗传相似系数计算、UPGMA方法聚类。
2 结果与分析
2.1马蹄金的RAPD和SRAP标记多态性分析 20份RAPD引物共扩增出189条带,其中130条具有多态性,多态性条带比率(PPB)为68.78%,扩增条带变幅6~13条,每个随机引物平均产生6.5条多态性条带(表2、图1)。多态性最高的引物是SB5,达到100%;多态性最低的引物是SB6,仅为30.00%。相比之与RAPD,22对SRAP引物共计扩增条带242条,其中多态性条带为175条,占总条带的72.31%,引物组合Me5+Em4扩增条带最多达14条,组合Me6+Em6和组合Me8+Em2扩增条带最少,仅为7条,每对引物平均扩增出7.95条多态性条带(表3)。引物组合Me5+Em9多态性高达到90.00%,组合Me8+Em9多态性最低,仅为58.33%。相比较而言,SRAP分子标记所检测到的马蹄金多态性略高于RAPD。
2.2马蹄金亲缘关系分析 利用NTSYS-pc软件,对20个RAPD引物的扩增结果进行遗传相似系数(Genetic Similarity,GS)计算。GS值变化范围为0.323~0.984,平均值为0.665,遗传距离在0.016~0.677,平均遗传距离为0.335。GS最大值同时出现在两组四川材料中,即材料SD200309与材料SD200310以及材料SD200308与材料SD200411之间,同为0.984;材料SD200303与材料SD200512遗传相似系数最小,仅为0.323,表明二者之间亲缘关系最远。SRAP标记下,GS值的变化范围在0.479~0.942之间,平均值为0.716,遗传距离在0.058~0.521,平均遗传距离为0.284。与RAPD结果不同,GS最大值出现在两份云南材料YD200505和YD200506之间,为0.942;GS最小值则同时出现在两份云南材料YD200504与YD200506,以及美国进口品种Common与云南材料YD200506之间,仅为为0.479。两种分子标记方法所检测到的马蹄金亲缘关系存在一定的差异。
2.3马蹄金RAPD聚类分析 根据遗传相似系数,21份供试马蹄金被划分为2个类群(图3)。结合王昆蕾等[21-22]对马蹄金形态特征的研究来看,聚类结果与其形态特征相关。第Ⅰ类群包括9份材料,这一类群种质叶片偏大,叶色深绿,除SD200303外,其他8份材料分枝能力较弱;SD200304、SD200303和CK在叶色、 叶片大小上都较为相似。第Ⅱ类群包括12份材料,这一类群种质叶片较第Ⅰ类小,叶色绿。

表2 20个RAPD引物对21份马蹄金材料扩增结果
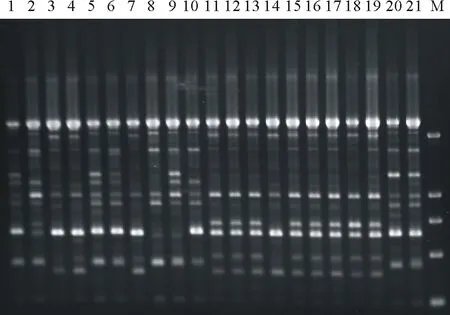
图1 引物SC6对21份马蹄金扩增的RAPD图谱

表3 22对SRAP引物对21份马蹄金材料的扩增结果
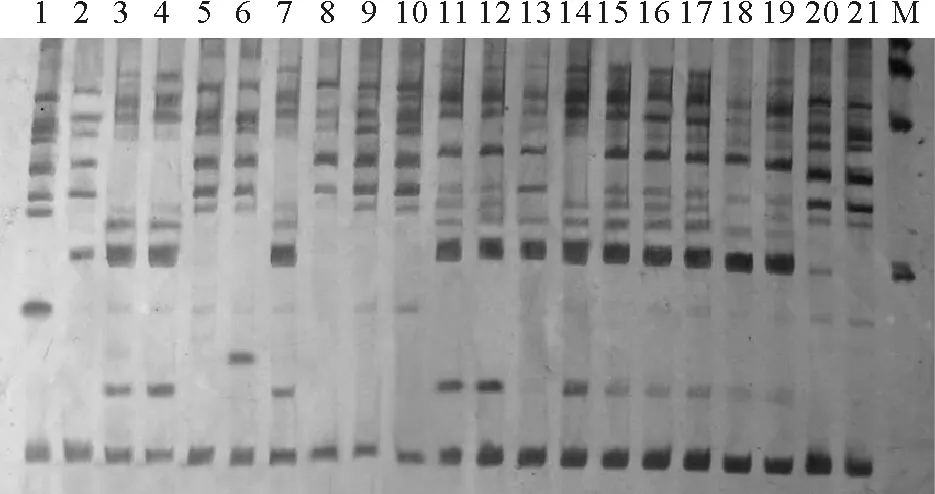
图2 Me2+Em10引物组合的SRAP 扩增谱带
2.4马蹄金SRAP聚类分析 21份供试材料同样被划分为2个大类(图4),该聚类结果与笔者选用19份SRAP引物所得的聚类结果比较相似[23-24]。第Ⅰ类群由2份云南材料YD200503、YD200504、5份四川材料SD200304、SD200303、SD200405、SD200801、SD200407、1份贵州材料GD200502以及美国品种Common组成。尽管美国品种与材料GD200502聚类结果较近,且同时划分在第Ⅰ类群,但根据笔者区域试验显示[23],Common这份材料绿色期、叶柄长度、抗性等多个坪用特性较之于其他材料有很大差异。第Ⅰ类群由2份云南材料YD200506、YD200505、8份四川材料SD200302、SD200309、SD200512、SD200406、SD200513、SD200310、SD200308、SD200411 和2份贵州材料GD200504、GD200503组成,该类群材料叶片较小,叶柄较短,叶色绿,绿期较长。由此可见,22份马蹄金材料中,除了贵州材料GD200502和美国进口品种Common的聚类没有明显的联系外,来自相似生态环境的材料能大致聚在一起,且聚类结果与形态学特征、坪用特性也表现出一定相关性。
2.5RAPD和SRAP相关性分析 本研究用筛选出的20个RAPD引物和22对SRAP引物组合对21份马蹄金材料进行PCR扩增,RAPD扩增结果的GS值在0.323~0.984,平均GS值为0.665。SRAP扩增结果的GS值在0.479~0.942,平均值为0.716,差别不大。根据所得条带进行聚类分析,将21份马蹄金材料按亲缘关系远近划分为不同的类群,RAPD标记和SRAP标记都将21分材料划分为2个类群,并可将SRAP聚类结果看作为RAPD聚类的细化。为了检测供试材料间的RAPD和SRAP分析结果的相关程度,利用NTSYS-pc 2.10软件的mantel对两组数据进行了相关性分析,相关系数r0.01=0.910,二者存在极显著正相关,说明优化后的RAPD和SRAP技术较为稳定,表明应用这2种技术对马蹄金遗传多样性与亲缘关系的分析具有较高的一致性和可信度。

图3 基于Nei-Li 相似系数的21份马蹄金材料RAPD-UPGMA聚类图
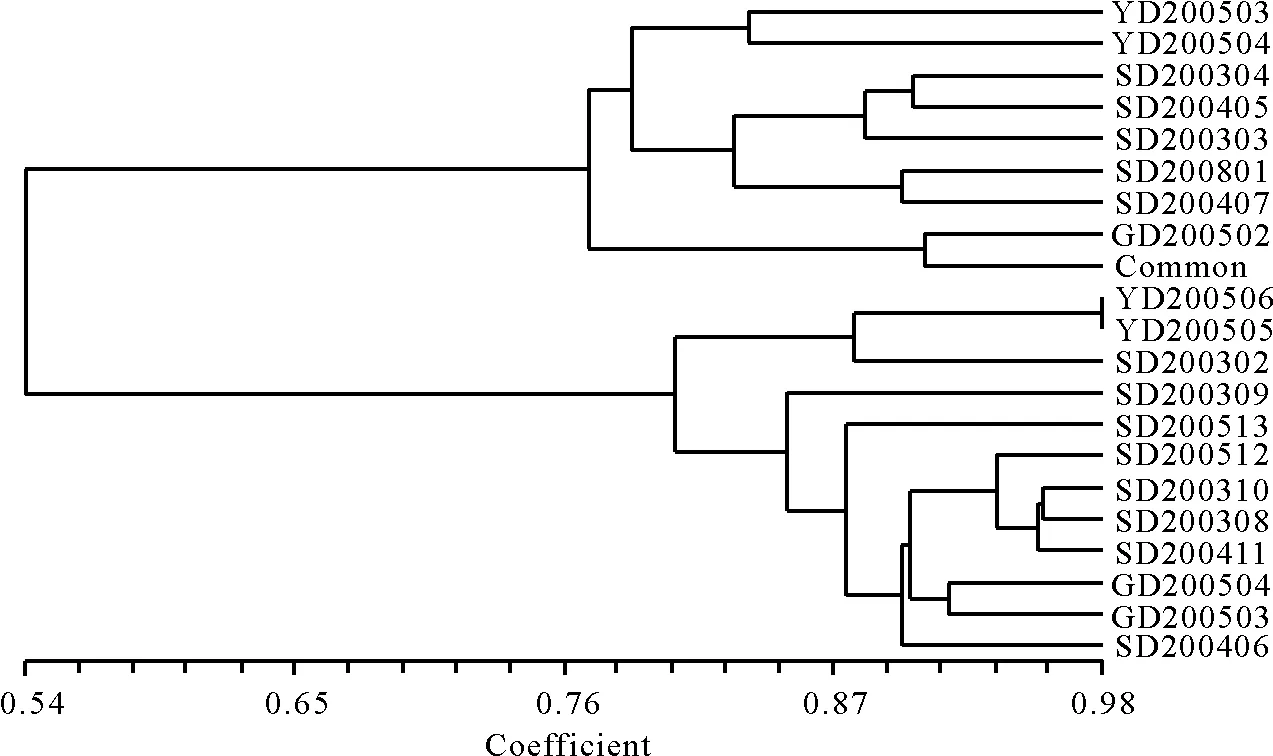
图4 基于Nei-Li 相似系数的21份马蹄金材料SRAP-UPGMA聚类图
3 讨论
3.1马蹄金种质资源的遗传多样性 王昆蕾等[21-22]对23份野生马蹄金材料进行了形态学性状和ISSR遗传多样性研究,结果表明,我国野生马蹄金在形态上存在着丰富的遗传变异,其中叶长、叶宽、草层高度、叶柄长、分枝数、主茎长以及叶片、匍匐茎颜色的变异都超过20%;形态学聚类分析将23份野生材料分为4个类群:大叶高丛型、大叶矮丛型、小叶矮丛型、小叶短茎型,ISSR中23份野生马蹄金材料GS值为0.211~0.871,平均GS值0.394,该分子标记方法检测的材料间遗传距离较近。本研究通过RAPD和SRAP分子标记技术对西南地区野生马蹄金植物的遗传多样性进行研究,从结果看,RAPD扩增结果的GS值在0.323~0.984,平均GS值为0.665,SRAP扩增结果的GS值在0.479~0.942,平均值为0.711,2种分子标记结果的遗传多态性分别为68.78%与72.31%,说明本研究所用的种质具有一定代表性,且这些结果从分子标记的角度进一步说明了我国西南地区野生马蹄金种质资源间存在丰富的遗传变异。
3.2聚类结果与形态学相关性 聚类结果与形态学具有一定的相关性。在RAPD聚类中,云南材料YD200503、YD200504、四川材料SD200304、SD200303、SD200405、SD200801、SD200407、贵州材料GD200502以及对照品种Common聚为第Ⅰ类,这9份材料的叶片偏大,草层高度整体较第Ⅱ类材料高,主茎长度总体比第Ⅱ类材料稍短;第Ⅱ类的12份种质除SD200308叶片相对较大以外,其他11份材料叶片较第Ⅰ类小,主茎节总体长于第Ⅰ类群,草层矮,分枝能力强。SRAP聚类与RAPD基本相同,材料的栽培品种,由于长期适应不同生长尽管贵州材料GD200502和美国对照品种的遗传相似系数达0.905,亲缘关系近,但两者在形态特征和坪用特征上差异较大,解释这一现象的最大可能性为GD200502是另一个有别于对照环境导致外观形态出现变异,对二者是否属于同一种可在今后的试验中进一步深入研究。此外,近几年来,为更好研究物种遗传多样性,研究者们倾向于采用多种分子标记探索物种基因信息。有研究[23]表明,不同的分子标记研究的结果可能是一致的,也可能并不一致,在许多情况下,形态同基因型间并无直接联系,形态性状的趋异不一定能反映遗传上的趋异,形态性状的相似也不一定意味着遗传关系的近缘,两个形态上有差异的个体可能在遗传上非常的相似[25]。
3.3聚类结果与地理分布相关性 聚类结果除了与形态特征具有一定相关性外,与马蹄金起源及分布范围有关,这一结论与王昆蕾[22]ISSR分析结论相似,即:相似形态特征的材料能聚成一类,相近生境条件的材料也能聚成一类。如来自云南的材料YD200503、YD200504、YD200506、YD200505,虽然没有聚在同一大类,但它们也在各大类中单独聚在一起。四川材料SD200304、SD200303、SD200405、SD200801、SD200407,这5份材料遗传距离较近,RAPD和SRAP聚类中,5份材料都能首先聚在一起,导致这种现象的原因可能与它们的起源和分布区域相似性有关。来自云南的2份材料YD200503和YD200504生态环境相似,也能相互聚在一起。另外,来自云南的2份材料YD200506、YD200505亲缘关系很近,在SRAP遗传相似系数分析中,二者GS值最高,达0.942,RAPD遗传相似系数分析中GS值也高达0.934,二份材料的生态环境较接近,初步猜测很有可能是同一种,但两者是否属同一材料还有待于进一步研究。
3.4西南区野生马蹄金种质资源的保护和开发利用 本研究结合前期形态遗传多样性研究,以1份美国品种为对照,对西南地区20份野生马蹄金材料进行了RAPD和SRAP分子遗传标记,通过两种分子标记方法对比实验,初步对不同野生马蹄金植物形态特征及生物学特征存在的差异进行研究,了解马蹄金种资源遗传变异及其地理分布规律,对今后种质的保护研究以及野生资源的有效合理利用有重要意义。研究结果表明,我国野生马蹄金材料间存在丰富的遗传变异,应加强对其原生境保护或异地保护,防止人类活动造成的基因混杂。我国有大量野生马蹄金分布,今后可扩大采集范围,进一步开展野生材料的收集工作,为国内优良本土草坪草及地被植物驯化和育种提供丰富的种质资源。
[1]谭继清,谭志坚.新编中国草坪与地被[M].重庆:重庆出版社,2000:46-49.
[2]谢彩云,尚以顺.优良草坪植物马蹄金的生物学特性及栽培管理技术[J].贵州农业科学,2001,29(1):51-52.
[3]张晓慧,干友民,任婷,等.5份西南野生马蹄金材料的光合特性比较[J].草业科学,2010,27(3):76-79.
[4]李君.野生和栽培马蹄金抗旱性比较及其抗旱机制初探[J].植物生态学报,2007,31(3):521-527.
[5]邱菊.西南区野生马蹄金抗旱性研究[D].雅安:四川农业大学,2009:38.
[6]干友民,任婷,陈燕,等.西南地区野生马蹄金无性繁殖特性研究[J].草业科学,2009,26(8):163-171.
[7]陈燕,刘忠义,干友民,等.野生马蹄金无性系构件种群组成及生物量结构变异性[J].江西农业学报,2008,20(2):23-25,31.
[8]陈燕.西南区野生马蹄金繁殖特性研究[D].雅安:四川农业大学,2008:27.
[9]王钦,干友民,杨啸枫,等.四川野生马蹄金坪用特性研究[J].安徽农业科学,2005,33(4):630-631.
[10]周莹洁,王显国,张新全.野牛草种质基于SRAP标记的遗传多样性[J].草业科学,2011,28(11):1930-1935.
[11]井赵斌,余靓,魏琳,等.本氏针茅SRAP-PCR反应体系的建立及引物筛选[J].草业科学,2012,29(2):219-228.
[12]Li G,Gao M,Yang B,etal.Gene for alignment between theBrassicaandArobidopsisgenomes by direct transcriptome mapping[J].Theoretical and Applied Genetics,2003,107:168-180.
[13]郭海林,刘建秀,高鹤,等.结缕草属优良品系SSR指纹图谱的构建[J].草业学报,2007,16(2):53-59.
[14]郭海林,郑轶琪,陈宣,等.结缕草属植物种间关系和遗传多样的SRAP标记分析[J].草业学报,2009,18(5):201-210.
[15]李杰勤,王丽华,詹秋文,等.高粱棕色中脉基因bmr-6的遗传分析和SSR标记定位[J].草业学报,2010,19(5):273-277.
[16]郑轶琦,刘建秀.草坪草分子遗传图谱的构建与应用研究进展[J].草业学报,2009,18(1):155-162.
[17]张西西,徐进,王涛,等.万寿菊杂交一代遗传多态性的SRAP标记分析[J].园艺学报,2008,35(8):1221-1226.
[18]田志宏,邱永福,严寒,等.用RAPD标记分析草地早熟禾遗传多样性[J].草地学报,2006,14(2):120-123.
[19]Li G,Quiros C F.Sequence-related amplified polymorphism(SRAP),a new marker system based on a simple PCR reaction:its application to mapping and gene tagging in Brassica[J].Theoretical and Applied Genetics,2001,103:455-461.
[20]袁学军,王志勇,郑轶琦,等.假俭草体细胞抗寒突变体的获得及其SRAP分子鉴定[J].草业学报,2011,20(6):237-244.
[21]王昆蕾,干友民,费凌,等.西南区野生马蹄金居群生态特性及形态变异[J].湖北农业科学,2006,45(6):798-798.
[22]王昆蕾.西南区野生马蹄金遗传多样性研究[D].雅安:四川农业大学,2007:41-42.
[23]付薇.野生马蹄金种质资源RAPD和SRAP分析及优异选系区域适应性研究[D].四川农业大学硕士学位论文.2010.
[24]付薇,干友民,吴佳海,等.野生马蹄金种质遗传多样性的SRAP研究[J].草业学报,2010,19(6):165-170.
[25]文雁成,王汉中,沈金雄,等.用SRAP标记分析中国甘蓝型油菜品种的遗传多样性和遗传基础[J].中国农业科学,2006,39(2):246-256.
