丝茅草侵入量与高羊茅相对竞争力及对幼坪性状影响的分析
2012-04-16刘金平游明鸿
刘金平,游明鸿
(1.西华师范大学生命科学院,四川 南充637009;2.四川省草原科学研究院,四川 成都611731)
*杂草不仅损害草坪的美观性、影响草坪草生长发育、增加草坪养护难度和强度,且杂草是诸多病虫害的中间寄主[1-3]。幼坪期草坪的密度低、盖度差、抗性弱,杂草极易入侵且只能人工拔除。防除杂草是幼坪养护最重要和最基本的工作[4],除草用工占总用工的80%左右,费用占管理费用的60%左右。开展入侵杂草对草坪草竞争压力的研究,对草坪养护、杂草防除及降低管理投入具有重要现实意义。
丝茅(Imperatakoenigii或I.cylindricavar.majorb)为禾本科白茅属植物,即俗称的茅针、茅根、白茅根,在全球热带和亚热带地区广泛分布[5],其根茎发达、种子繁多,常侵占土地和森林,毁坏农作物,破坏本土植物,导致土质下降,颠覆生态系统,被认为是世界上最恶毒最完美的10种杂草之一[6]。围绕其侵入、扩散及防除进行了大量研究[7-9],但目前仍无有效、持久的防除措施。在亚热带及过渡性气候带,丝茅是冷季型草坪最常见恶性杂草,一旦入侵很难剔除。入侵1~3年后,常在草坪中形成局部优势种群斑块,甚至完全代替草坪草,严重危害草坪种群结构与景观功能。
本试验以冷季型草种中抗逆性极强的高羊茅(Festucaarundinacea)为材料,模拟丝茅草在建植时入侵,通过测定相对产量、相对产量总和、攻击指数、株高、分蘖数等指数,分析丝茅草侵入量对高羊茅竞争力、草坪性状及草坪品质的影响,探讨抑制丝茅草危害、提高草坪品质的有效途径,为草坪建植、养护、管理提供依据。
1 材料与方法
1.1 试验材料及设计
高羊茅为丹麦生产的黄金岛(Eldorado)品种,丝茅草为当地采集的野生种子。采用高34 cm、口径50 cm塑料花盆,河沙腐殖土1∶1为基质。
参考朱宏伟等[10]方法进行取代试验设计。2010年3月,按单播和混播2种方式建立模拟混生草坪种群。单播时高羊茅和丝茅草分别按50株/盆进行定苗,设3次重复。混播时按高羊茅和丝茅草9∶1,8∶2,7∶3,6∶4,5∶5比例,50株/盆,均匀分布进行定苗,均设3次重复。在25℃左右的室内培养。期间正常浇水、除杂,于20,40,60 d进行3次留茬高度9 cm的修剪,70 d时进行测定。
1.2 测定项目与方法
草坪品质:第70天,以密度、颜色、质地和均一性为评分指标,采用九分制标准进行评定[11,12]。
株高、分蘖与根系深度:第70天,随机选取高羊茅、丝茅草各10株(不足10株时,全样法),测定单株自然高度、分蘖数,而后倾倒出盆,剔除泥土后,测定根系深度(80%根达到的深度)。
生物量:第70天,倒出盆后,小心分离,随机选取高羊茅、丝茅草各10株,分离地上与地下部分,清洗分别装袋,105℃下烘至恒重后称重,计算单株与每盆的地上、地下生物量。
相对产量 (relative yield,RY):按照 Fowler[13]的公式进行:RYij=Yij/(p Yi)和RYji=Y ji/(qYj)计算。RYij代表与种j混播时种i的相对产量,RYji代表播时种i的相对产量。Yij是与种j混播时种i的生物量,Y ji是播时种j的生物量,Y i是单播i的生物量,Y j是单播j的生物量。生物量均以盆为单位,而相对产量则以株为单位。p是混播方式下种i的比例,q为j的比例。当RY<1.0时,表示种间竞争大于种内竞争;当RY>1.0时,种内竞争大于种间竞争;而当RY=1.0时,表明种内和种间竞争水平相当。
相对产量总和 (total relative yield,RYT)按公式:RYT=p RYij+qRYji进行计算。RYT<1.0表明两物种间有拮抗作用,RYT>1.0表明两物种之间没有竞争,RYT=1.0表明两物种需要相同的资源,且一种可通过竞争将另一种排除出去。
攻击力指数 (attack index,A):根据 Mc Gilchrist和Trenbath[14]的公式:Ai=RYij-RYji和A j=RYji-RYij计算。
1.3 数据分析
用SAS 9.1统计分析数据。
2 结果与分析
2.1 丝茅草侵入量与高羊茅相对竞争力
2.1.1 相对产量(RY) 丝茅草入侵使高羊茅茎叶、根系受到的种间竞争大于种内竞争,高羊茅地上、地下相对产量均小于1.0。高羊茅对丝茅草的种间竞争小于其种内竞争,丝茅草地上、地下相对产量均大于1.0。多重比较说明,丝茅草侵入量对高羊茅相对产量有极显著影响(P<0.01)(表1),侵入量越大对高羊茅茎叶竞争胁迫越显著(P<0.05),对根系胁迫小于茎叶。丝茅草侵入量50%时其地上RY才显著受种内竞争的影响(P<0.05)。

表1 丝茅草侵入量与高羊茅相对竞争力的多重比较Table 1 Multiple comparisons about intrusion volume of Imperata and relative competitiveness of Festuca
2.1.2 相对产量总和(RYT) 丝茅草侵入量对群落的地上、地下相对产量总和有极显著影响(P<0.01)(表1)。地上、地下RYT的值均小于1.0,表明两物种间有拮抗作用。地上RYT值在丝茅草侵入量为50%时最小,群落中地上茎叶的种间、种内竞争合力最强。地下RYT值则在侵入量为30%时最小,说明两物种间地上、地下拮抗作用不存在协同性。当F∶I=9∶1时,丝茅草影响小,群落产量以高羊茅为主。F∶I=8∶2~6∶4时2种植物竞争激烈,群落产量由两者共同决定,地上RYT值相对平稳,而地下RYT值以F∶I=7∶3作为拐点急剧升降。当F∶I=5∶5时,高羊茅受抑制,群落以丝茅草为主。
2.1.3 竞争攻击力(A) 丝茅草侵入量对丝茅草的地上、地下攻击力指数有极显著影响(P<0.01)(表1)。丝茅草侵入量越大A值越大,其在群落中的竞争力越强。高羊茅竞争攻击力及攻击指数与丝茅草相反。随侵入量增加,丝茅草种间竞争增加,丝茅草地上攻击指数上升斜率大于地下。当F∶I=6∶4~5∶5时,丝茅草茎叶种内竞争加强,地上A值相对变平缓,而丝茅草根系在种间竞争中取得优势,地下A值快速增加。
2.2 丝茅草侵入量对高羊茅幼坪性状的影响
2.2.1 侵入量对高羊茅单株生物量的影响 混播下丝茅草单株的地上、地下生物量显著高于单播(P<0.05)(表2),当侵入量大于10%时,受其竞争力影响,高羊茅单株地上生物量极显著下降(P<0.01),侵入量越大差异越显著(P<0.05)。侵入量对地下生物量有极显著影响(P<0.01),当侵入量大于30%时,地下生物量受丝茅草竞争影响而显著降低。
2.2.2 侵入量对高羊茅单株分蘖数的影响 混播与单播对丝茅草单株分蘖数影响较小(P>0.05),随侵入量增加密度增加,丝茅草对地表空间的强竞争力,致使高羊茅单株分蘖数存在显著差异(P<0.05)。当侵入量大于10%时,高羊茅分蘖数显著低于单播,不同混播比例间有较大差异。当侵入量大于40%,其分蘖能力下降43%。
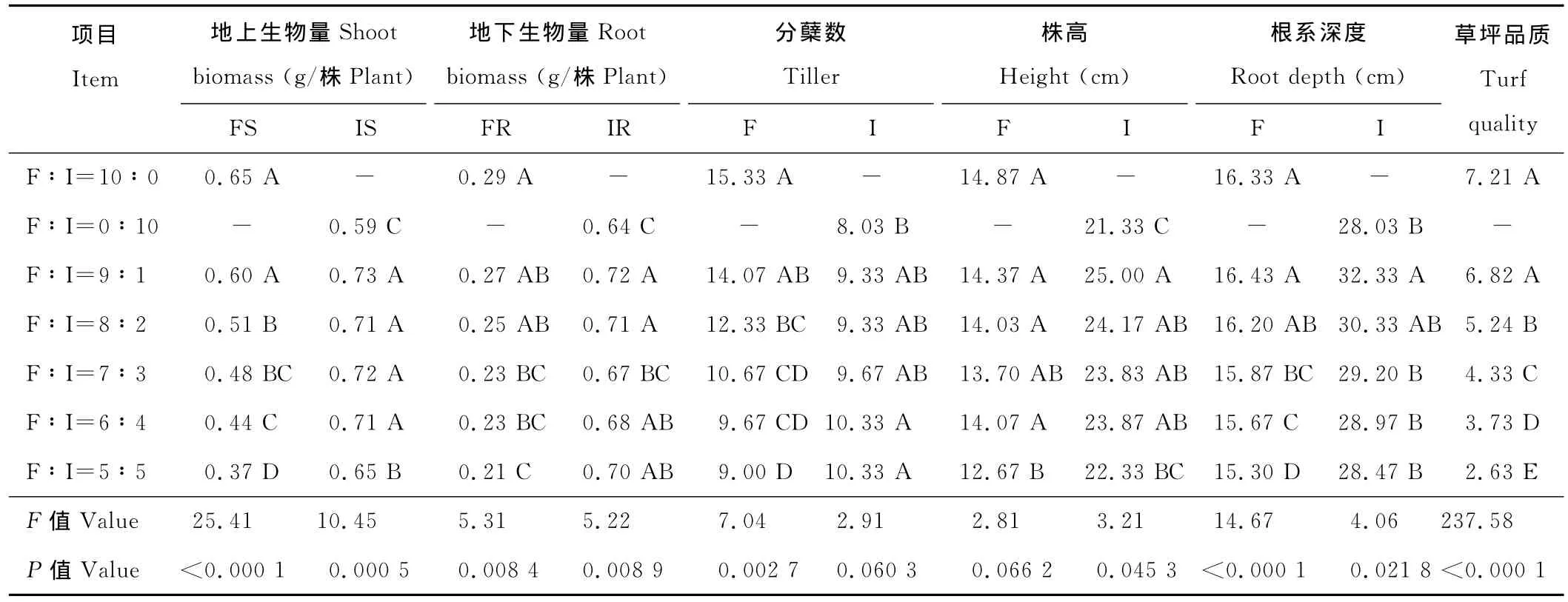
表2 丝茅草侵入量对高羊茅幼坪性状的影响Table 2 Effect of intrusion volume of Imperata on young turf traits of Festuca
2.2.3 侵入量对高羊茅株高的影响 混播下丝茅草的高度显著高于单播(P<0.05)(表2),随侵入量增大,丝茅草密度增加,种内竞争使其高度下降。丝茅草高度与密度对共生高羊茅密度(分蘖数)影响显著,但对株高影响较小,仅当侵入量50%时,株高才显著下降。
2.2.4 侵入量对高羊茅根系深度的影响 丝茅草根系深于高羊茅,低侵入量下由于种内竞争小,混播丝茅草根系深于单播。随侵入量增加,丝茅草根系深度变化不大,但种间竞争压力使高羊茅根系显著受到影响,当侵入量大于30%后,高羊茅根系显著变浅。不同侵入量对高羊茅根系深度有极显著影响(P<0.01)。
2.2.5 侵入量对高羊茅草坪品质的影响 丝茅草对株高、分蘖的影响,使草坪密度、均一性下降,2种植物生物学特性不同使草坪颜色、质地差异增大。侵入量对草坪品质有极显著影响(P<0.01),随丝茅草比例增加,草坪品质显著下降。
2.3 丝茅草侵入量与高羊茅竞争指数及性状指数的相关分析
丝茅草侵入量除与高羊茅株高相关性不显著外(表3),与高羊茅竞争力指数(相对产量、攻击指数)呈极显著负相关,与高羊茅分生再生能力(分蘖)与抗逆性(根系深度)指数呈极显著负相关,与草坪品质指数呈极显著负相关。可见,丝茅草入侵对高羊茅草坪的影响有持续性与系统性。
高羊茅地上攻击指数(X4)与地上生物量、地下生物量、分蘖数、根系深度呈显著正相关(表3),而与株高相关性较小。同理,地下地上攻击指数(X4)与上述参数均为显著正相关。可见,促进分蘖、增加密度、加快生物量积累是提高其竞争力的有效途径。
草坪品质(X11)与丝茅草侵入量呈极显著负相关,与株高外的其他指数呈显著正相关(表3)。可见,防除杂草、抑制株高、促进分生是提高草坪品质的基本要求。

表3 丝茅草侵入量与高羊茅竞争指数及性状指数的相关分析Table 3 Correlation analysis about intrusion volume of Imperata and competition index,traits index of Festuca
3 讨论与结论
研究混生种群的竞争行为,常采用部分增加、取代系列、添加系列、完整添加4种基本设计方式,每种设计都有相关的优缺点[15]。本试验采用复合de Wit取代设计,保持混生种群总密度不变,只改变种群的组成比例。取代系列设计通常以植物产量(地上生物量)为核心内容,用相对产量、相对产量总和及攻击指数来研究竞争关系。考虑到草坪地被结构、功能的特殊性,本试验采用地上、地下生物量,从立体角度分析两者的竞争情况。高羊茅地上、地下RY均小于1.0,以种间竞争为主,丝茅草均大于1.0,以种内竞争为主。随入侵比例增加,丝茅草种内竞争逐渐下降,整个群落种间竞争加强。混生种群地上、地下RYT的值均小于1.0,表明两物种间有拮抗作用,但地下RYT均近似于1.0,说明这2种植物根系存在着对相同资源的直接竞争。同时丝茅草攻击力为正,高羊茅为负,前者的地上、地下竞争力均大于后者,随入侵比例增加,其竞争优势越明显。对相同资源的直接竞争及拮抗作用,必然使群落格局与其优势种的格局存在潜在的定向性演替风险。群落组成及分布格局不仅受优势种的生物学特性影响,还受环境因子制约[16],营造适合草坪草生长的环境因子,是草坪建植与养护的技术核心要求。修剪、灌溉、表施土壤等养护措施,实质是为草坪草竞争力提高而提供动力。所以,加强养护管理是防止丝茅草入侵与泛滥的有效途径。
本研究模拟草坪建植时丝茅草入侵,对幼坪期入侵物种与目标植物的竞争进行了分析。竞争结果最终通过植物生长直接表现,包括植株高低、叶片宽窄、茎粗细等现实表现,分蘖等分生再生能力和种子产量等生育力所蕴含的潜在表现。竞争植物的现实表现和潜在表现差异,决定其在时间空间延展过程中,适合度、优势度、存活率与净繁殖率等特征的发展趋势,使植物群落的组成、结构与功能发生改变。另外,群落植物竞争是全方位与连续的过程,用某个时间点植物的表现与密度来表达彼此的竞争力,其实表述的只是以前时间段累积的竞争结果,而不能具体说明现在的竞争强度及将来的竞争趋向。尤其丝茅草为根茎型禾草,其庞大的根系系统与快速生长能力,使其在地上、地下资源竞争中占有现实优势,其强大的根茎分生、再生潜力与超强的种子繁殖能力,使其在将来种群组成比例上占有潜在优势。并且丝茅草根茎含有醇、萜醇、甾醇、酸类等[17]次生代谢产物,通过影响土壤理化性质,必然对其共生植物有一定的化感作用。入侵丝茅草对高羊茅的竞争是立体而系统的,在时间、空间与环境因子的契合情况下,其潜在优势将转化为现实优势,也将孕育更大更持久的潜在优势。根茎型禾草的株丛结构、分蘖株龄级组成及根茎生产力,随生长年限而发生变化[18],变化程度直接决定其竞争力的大小。所以,有必要开展深入系统的研究,分析丝茅草对冷季型草坪竞争的动态变化,探讨其危害发生、发展规律,为该类恶性杂草的防除提供依据,为草坪建植养护提供全面、深入、持续的服务。
试验以密度、颜色、质地和均一性等外观指标,对草坪品质进行评价。丝茅草叶片宽大、叶脉发达、颜色灰绿,分蘖能力、生长速度明显高于高羊茅,丝茅草占种群地上生物量的比例及高度直接决定草坪的外观质量。修剪可以破坏顶端优势,促进分蘖的形成,是提高草坪品质的基本养护措施[19,20]。试验模拟冷季型草坪的修剪频度,每隔20 d修剪1次。结果说明在正常养护水平下,根茎-丛生型丝茅草入侵量越大,丛生型高羊茅单株分蘖数及种群密度比例越小。丝茅草发达的根茎与深根系,使其在地下空间、水分、营养的竞争中占有优势,致使其生长速度明显高于高羊茅。入侵植物粗糙的形态特征与增加的群落比例,使草坪品质随其侵入量增大而降低。草坪品质与侵入量呈极显著负相关,与株高外的其他指数呈显著正相关。群落内物种间的竞争强度,不仅决定植物将能量用于生长或竞争,并且与植物保护酶含量及其抗性有关[21]。竞争中高羊茅的能量主要用于竞争,致使分蘖等生长受到限制,最终导致了草坪功能的退化。可见,防除杂草、抑制株高、促进分生,是提高羊茅草坪品质的基本要求与必然途径。
本试验以冷季型草坪草中适应性、抗逆性最强的高羊茅为材料,说明高羊茅与丝茅草竞争中处于劣势,竞争结果将是前者逐渐被后者排挤出草坪,致使草坪结构与功能彻底丧失。过渡性气候带地区,常以高羊茅为建群种、草地早熟禾(Poapratensis)为伴生种、多年生黑麦草(Loliumperenne)为保护种,来混播建植冷季型庭园草坪,经2年左右后,往往成为高羊茅单一草坪。本试验仅就丝茅草侵入量对幼坪期草坪草的竞争力、单株生长及草坪品质的影响进行了分析。由于冷季型草种自身生物学特点、生态学特性的局限性(如夏眠、丛生型、分生再生能力等)、养护技术不合理与养护投入低等原因,随时存在丝茅草入侵的风险。所以,开展丝茅草繁殖特点、入侵途径及对冷季型草坪危害规律的深入研究,是防除恶性杂草,提高草坪品质、延长草坪寿命的必然要求。
[1] 谭永钦,张国安,郭尔祥.草坪杂草生态位研究[J].生态学报,2004,24(6):1300-1305.
[2] 强胜,李广英.南京市草坪夏季杂草分布特点及防除措施研究[J].草业学报,2000,9(1):48-54.
[3] 吴海荣,强胜.南京市秋季外来杂草定量调查研究[J].生物多样性,2003,11(5):432-438.
[4] 刘金平,毛凯,游明鸿.草坪在生态城市建设中的作用及其应用[J].草业科学,2002,19(2):50-53.
[5] 冷琴,杨雅玲.国产白茅属部分生物学特性的研究[J].南京大学学报(自然科学版),2002,38(5):703-714.
[6] 徐立荣.草坪杂草——白茅防治技术[J].南方农业,2008,2(10):76-78.
[7] Willard T R,Shilling D G,Gaffney J F,etal.Mechanical and chemical control of cogongrass(Imperatacylindrica)[J].Weed Technology,1996,10:722-726.
[8] King S E,Grace J B.The effects of gap size and disturbance type on invasion of wet pine savanna by cogongrass,Imperata cylindrica(Poaceae)[J].American Journal of Botany,2000,87:1279-1286.
[9] Otsamo R.Early development of three planted indigenous tree species and natural under storey vegetation in artificial gaps in anAcaciamangiumstand on anImperatacylindricagrassland site in South Kalimantan,Indonesia[J].New Forest,2000,19:51-68.
[10] 朱宏伟,孟玲,李保平.黑麦草与入侵杂草紫茎泽兰苗期的相对竞争力[J].应用与环境生物学报,2007,13(1):29-32.
[11] 刘金平,游明鸿,毛凯,等.不同季节叶施N、Fe对假俭草草坪品质的影响[J].四川草原,2004,(3):50-52.
[12] 刘金平,游明鸿,毛凯.秋冬季叶施N、Fe提高假俭草抗寒性的研究[J].亚热带植物科学,2004,33(1):12-15.
[13] Fowler N.Competition and coexistance in a north carolina grassland[J].Journal of Ecology,1982,70:77-92.
[14] Mc Gilchrist C A,Trenbath B R.A revised analysis of plant competition experiments[J].Biometrics,1971,27:659-671.
[15] Jonathan S,Deborah C.简明植物种群生物学[M].李博,董惠琴,陆建中,等译.北京:高等教育出版社(第四版),2003:175-178.
[16] 李素清,武冬梅,王涛,等.山西长治湿地草本植物优势种群和群落的空间格局分析[J].草业学报,2011,20(3):43-50.
[17] 丘丹萍,邹勇芳,黄锁义.白茅根多糖提取方法的比较研究[J].中国酿造,2010,1:108-109.
[18] 刘金平,毛凯,游明鸿.假俭草草坪管理技术研究[J].四川草原,2001,(4):37-40.
[19] 江海东,曹卫星,王举斌.修剪对高羊茅生长及草坪质量的影响[J].草业科学,1998,(2):54-61.
[20] 李海燕,李建东,徐振国,等.内蒙古图牧吉自然保护区羊草种群营养繁殖特性的比较[J].草业学报,2011,20(5):19-25.
[21] 王金龙,赵念席,徐华,等.不同地理种群大针茅生理生化特征的研究[J].草业学报,2011,20(5):42-48.
