补播对高寒草甸生产力和植物多样性的影响
2012-04-12张永超牛得草韩潼陈鸿洋傅华
张永超,牛得草,韩潼,陈鸿洋,傅华
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020)
*青藏高原由于其独特的自然环境,作为气候变化的敏感区域,其脆弱生态系统的发生发展,成为人们关注的热点之一[1]。自20世纪80年代以来,青藏高原草地生态系统严重退化,局部地区向“黑土滩”和沙漠化方向发展,影响了当地畜牧业的发展[2,3]。位于黄河第一弯的甘肃玛曲县,是黄河径流重要的汇集区和黄河上游至源头的重要水源涵养区[4,5],同时也是黄河上游生态系统的绿色生态屏障,是整个青藏高原生产力最高和生物多样性最为丰富的草地,素有“中国西部高原之肾”、“亚洲第一牧场”和“高原水塔”之称[6-10]。该区天然草地面积占土地总面积的84.3%,其中可利用草地面积占96.7%[11],丰富的资源为玛曲县经济的发展提供了得天独厚的条件。但近年来,在经济利益的驱使下,牧民超载过牧严重,导致自然生态系统严重失衡,生物多样性降低,草地生产力严重下降,草层高度降低,毒杂草比例上升,裸地增加,土壤蓄水保肥能力减弱,水土流失加剧,全县近90%的天然草地出现不同程度的退化和沙化[2,3,12]。因此,在该地区进行退化草地的恢复治理,对保护当地生态环境、增加草地涵养水源的功能,提高牧草产量促进当地畜牧业发展具有重要意义。
补播是退化草地恢复治理的有效途径之一,退化草地补播能大幅度提高草地生产力,增加优质牧草比例和植被盖度。据报道[13-16],在天然草地播种以沙打旺(Astragalusspp.)为主的优良牧草,草地产草量比对照提高4~5倍,同时补播豆科牧草沙打旺和禾本科牧草垂穗披碱草(Elymusnutans),能显著提高产草量,增加优质牧草比例,草层高度和密度显著提高。郑华平等[6]指出补播对高寒沙化草地物种丰富度影响不明显,但草群密度和地上生物量比对照显著增加。为从根本上遏制高寒草甸沙化,需在高寒草地退化初期采取恢复措施,然而关于高寒草甸退化初期补播改良的研究报道较少[6,17],本研究初步探讨了补播对高寒草甸退化草地恢复过程中群落生产力、功能群结构和物种多样性方面的影响,旨在为玛曲退化高寒草甸的改良恢复,牧区经济环境可持续发展提供技术指导和理论依据。
1 材料与方法
1.1 研究区自然概况
研究区位于青藏高原东部的甘肃省玛曲县阿孜畜牧科技园区,地处北纬33°40′,东经101°53′,海拔3 585 m。该区属于高原大陆性气候,年均温度1.2℃,1月平均温度-10℃,7月平均温度11.7℃。降水主要集中于5-9月,年降水量为564 mm,年蒸发量1 000~1 500 mm。全年日照时数2 613.9 h,一年的霜期大于270 d,没有绝对的无霜期[15,16]。土壤为亚高山草甸土。植被类型为高寒草甸草地,植物以莎草科的嵩草属(Kobresia)和禾本科的针茅属(Stipa)、羊茅属(Festuca)、早熟禾属(Poa)的一些种,以及菊科的凤毛菊属(Saussurea)、橐吾属(Ligularia)、瑞香科的狼毒属(Stellera)等毒杂草为主,并伴有其他杂类草,如高山唐松草(Thalictrumalpinum)、莓叶委陵菜(Potentillafragarioides)、钝裂银莲花(Anemoneobtusiloba)、细叶亚菊(Ajaniatenuifolia)、火绒草(Leontopodiumhastioides)、花苜蓿(Medicagoruthenica)等[6,7]。
1.2 试验设计
在研究区选择退化程度均一、地势平坦的退化草地,于2010年5月进行围封。试验设置4个处理,即不补播(B0),补播7.5 kg/hm2(B1)、15 kg/hm2(B2)、22.5 kg/hm2(B3),补播草种为垂穗披碱草。每个处理设5个重复小区,每个小区面积为15 m×10 m。2010年5月中旬将种子撒到小区,为提高补播效果,各处理在补播后统一进行划破措施。
1.3 观测项目和测定方法
于2010年8月中旬,在不同处理的小区内随机设置1个0.5 m×0.5 m的样方,进行群落学调查,记录每个样方内的植物种类、密度、株高、盖度,并将样方内的植物齐地面剪下,分种装入信封袋,实验室内105℃杀青30 min,65℃温度下烘干至恒重,计算生物量。
植物多样性分析采用Simpson多样性指数,Shannon-Weiner多样性指数,Margalef丰富度和Pielou均匀度指数进行多样性的测定,其计算公式如下:

式中,S为0.25 m2样方内的物种数,N为样方中物种总个体数,Pi为样方中第i种的密度在群落总密度中所占的比例。
1.4 统计分析
采用Excel进行制图,用SPSS 17.0进行方差分析。
2 结果与分析
2.1 补播对地上总生物量和功能群生物量的影响
相对于不补播处理(B0),补播使得地上总生物量显著提高1.31~1.44倍(图1),但各补播处理之间(B1、B2和B3)差异不显著。植物不同功能群间的比较表明,莎草类地上生物量随补播量的增加呈增长趋势,但处理间差异不显著;禾草类生物量在补播梯度B1(7.5 kg/hm2)处理下显著高于对照(B0),其他处理差异不显著;杂类草各处理间差异不显著。
2.2 补播对植物群落功能群结构的影响
植物群落功能群结构在不同补播处理间差异显著(图2),补播处理降低了杂类草在群落中的比例,其中B2和B3处理较对照差异显著。莎草类和禾草类在补播处理下较对照有所增加,但差异不显著。不同播量之间,莎草类、禾草类和杂草类比例差异不显著。
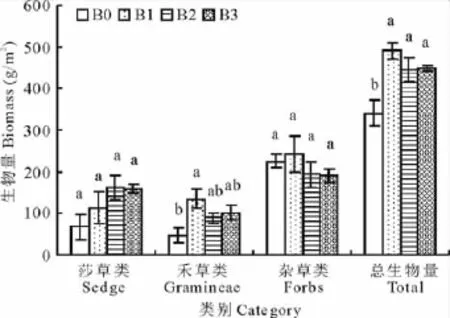
图1 不同补播梯度对退化高寒草甸草地地上总生物量和植物功能群生物量的影响Fig.1 Effect of different reseeding gradient on above-ground biomass and functional group productivity
2.3 补播对植物群落物种多样性的影响
相对不补播处理(B0),补播使得群落物种数(S)显著增加,而不同补播量之间没有显著差异。Margalef丰富度指数补播处理为4.051~4.127,显著高于对照(B0,2.952)。但Pielou均一性指数和Simpson及Shannon-Weiner多样性指数在各处理间没有显著性差异。
3 讨论与结论
研究表明高寒退化草地补播后地上总生物量显著提高(图1),此结果与与张云等[17],马玉明[18]和陈广夫等[19]的研究结论一致,可能是由于新物种的添加填充了原有退化草地空缺的生态位,新添加的物种生物量提高的缘故。不过,草地地上总生物量并没有随补播量的增加而显著提高,即不同播量之间,差异不显著,这可能是由于该实验于补播当年,新添加的物种还未在该生境中完全定居,很多种子还处于未萌发状态,补播效应还没完全显示出来,另一种可能是由于补播当年的效应,仅是表现在新添加的物种对原有退化草地空缺的生态位的填充,而随着年限的延长,高补播量带来的新添加物种在群落中的竞争优势可能才会得到体现[20]。此实验中所添加物种为禾本科植物垂穗披碱草,然而,对于植物功能群结构研究,并未发现各补播处理间禾草类植物地上生物量及其在群落中的比例显著增加。

图2 不同补播梯度对退化高寒草甸草地植物群落功能群结构的影响Fig.2 Effect of different reseeding gradient to the structure of functional group

表1 不同补播梯度对退化高寒草甸草地植物群落物种多样性的影响Table 1 Effect of different reseeding gradient on plant diversity
退化草地补播垂穗披碱草后,在补播当年尽管对莎草类植物地上生物量及其在群落中的比例没有造成显著影响,但是随着补播量的增加,莎草类绝对生物量及其在群落中的比例呈增加趋势。杂草类在中度和重度补播的条件下植物地上生物量显著下降,杂草类生物量及其在群落中的比例表现出逐渐降低的趋势,这可能是由于补播禾草后禾草类植物对土壤中养分利用的竞争,有效的抑制了杂类草的生长,而对刚刚引入系统的禾草类植物垂穗披碱草,此时还没能占据优势地位,草地系统依然以莎草类为主,而对杂类草植物抑制,同时,又为莎草类的扩展提供了新的生态位空隙,导致其生物量提高[21,22]。有试验报道[23],在高寒草地退化逐渐加剧的过程中,禾草类逐渐降低,而莎草类呈逐渐增加的趋势,可见适应于该区气候的顶级群落优势植物莎草类相对于禾草类有较强的竞争优势。
草地补播后,植物群落物种数(S)和丰富度指数(Margalef)较对照显著提高,这与Foster和Tilman[34]的研究结果一致,但是均匀度指数(Pielou)在各处理之间没有显著差异。可能是由于新的物种添加改变了原有群落物种竞争格局,有利于新的生态位空间的形成,导致群落物种数和丰富度增加[21,35]。而对于群落物种均匀度的提高,则需要长时间的物种竞争,重新调整格局,而对于补播当年,此种措施对群落物种均匀度影响还没表现出来。本实验Simpson多样性指数和Shannon-Weiner多样性指数在各个处理之间变化也不显著,可能主要受群落物种均匀度的影响。本研究结果发现,补播后植物群落物种数显著提高,表明群落中物种数是影响群落稳定性的主要因素,并且群落中物种越丰富其结构就越稳定[24-33],但鉴于垂穗披碱草为多年生牧草,补播垂穗披碱草对群落的稳定性的影响仍需要长期深入的研究。
致谢:感谢张卫国老师在前期样地设置和植物分类方面给予的帮助。
[1] 王素萍,宋连春,韩永翔,等.玛曲气候变化对生态环境的影响[J].冰川冻土,2006,28(4):556-561.
[2] 王向涛,张世虎,陈懂懂,等.不同放牧强度下高寒草甸植被特征和土壤养分变化研究[J].草地学报,2010,18(4):510-516.
[3] 曹建军,任正炜,杨勇,等.玛曲草地生态系统恢复成本条件价值评估[J].生态学报,2008,28(4):1872-1880.
[4] 王红梅.玛曲县草原荒漠化现状、成因及整治措施[J].草业科学,2005,22(11):20-24.
[5] 仁青吉,武高林,任国华.放牧强度对青藏高原东部高寒草甸植物群落特征的影响[J].草业学报,2009,18(5):256-261.
[6] 郑华平,陈子萱,牛俊义,等.补播禾草对玛曲高寒沙化草地植物多样性和生产力的影响[J].草业学报,2009,18(3):28-33.
[7] 马妙君,周显辉,吕正文,等.青藏高原东缘封育和退化高寒草甸种子库差异[J].生态学报,2009,29(7):3658-3664.
[8] 郭淑青,齐威,王玉林,等.青藏高原东缘海拔对植物种子大小的影响[J].草业学报,2010,19(1):50-58.
[9] 王娟,马文俊,陈文业.黄河首曲-玛曲高寒湿地生态系统服务功能价值估算[J].草业科学,2010,27(1):25-30.
[10] 曹广民,林丽,张法伟,等.青藏高原高寒矮嵩草草甸稳定性的维持、丧失与恢复[J].草业科学,2010,27(8):34-38.
[11] 朱丽,李广宇,魏强,等.玛曲退化草地生态修复研究[J].中国水土保持,2010,(1):28-38.
[12] 韩海涛,祝小妮.气候变化与人类活动对玛曲地区生态环境的影响[J].中国沙漠,2007,27(4):608-613.
[13] 卜繁超,姚文军.天然草地机械化补播改良技术及配套机具[J].新疆农机化,2002,(5):24-25.
[14] 安源,王育青,陈敏,等.沙地草场补播技术及其生态效益研究[J].草地学报,1997,(1):33.
[15] 陈文业,郑华平,戚登臣,等.黄河首曲沙化草地恢复重建模式研究[J].草业科学,2008,25(6):14-18.
[16] 王辉,任继周,袁宏波.黄河源区天然草地沙化机理分析研究[J].草业学报,2006,15(6):19-25.
[17] 张云,武高林,任国华,等.封育后补播“高寒1号”生态草对玛曲退化高寒草甸生产力的影响[J].草业科学,2009,26(7):99-104.
[18] 马玉明.碌曲县天然草地围栏封育及补播改良效果分析[J].现代农业科技,2009,(15):328-330.
[19] 陈广夫,蒋立宏,蒋景纯,等.鄂温克可旗带状补播改良退化草甸草原技术试验研究[J].内蒙古草业,2005,17(4):63-64.
[20] Tilman D.Niche tradeoffs,neutrality,and community structure:A stochastic theory of resource competition,invasion,and community assembly[J].Proceedings of the National Academy of Sciences,2004,101:10854-10861.
[21] Rohde K.Vacant niches and the possible operation of natural laws in ecosystems[J].Rivista di Biologia:Biology Forum,2008,101:13-28.
[22] Kawata M.Invasion of vacant niches and subsequent sympatric speciation[J].Proceeding of the Royal Society,2002,269:55-63.
[23] 陈文业,郑华平,戚登臣,等.甘南玛曲高寒草甸草地沙化的研究[J].草原与草坪,2008,127(2):77-80.
[24] May R M.Stability and Complexity in Model Ecosystems[M].Princeton:Princeton University Press,1973.
[25] Goodman D.The theory of diversity-stability relationships in ecology[J].Quarterly Review of Biology,1975,50:237-266.
[26] Mc Naughton S J.Diversity and stability of ecological communities:a comment on the role of empiricism in ecology[J].American Naturalist,1977,111:515-525.
[27] Pimm S L.The complexity and stability of ecosystems[J].Nature,1984,307:321-326.
[28] Tilman D.Biodiversity:population versus ecosystem stability[J].Ecology,1996,77:350-363.
[29] Hubbel S P.The Unified Neutral Theory of Biodiversity and Biogeography[M].Princeton:Princeton University Press,2001:1-151.
[30] Caswell H.Community structure:a neutral model analysis[J].Ecological Monographs,1976,46:327-354.
[31] Mc Kane A J,Alonso D,Sole R V.A mean-field stochastic theory for species-rich assembled communities[J].Physical Review,2000,62:8466-8484.
[32] Mc Kane A J,Alonso A D,Sole R V.Analytic solution of Hubbell’s model of local community dynamics[J].Theoretical Population Biology,2004,65:67-73.
[33] Wang Z K,Yang X Q.Birth-death Processes and Markov Chain(2nd ed.)[M].Beijing:Science Press,2005.
[34] Foster B L,Tilman D.Seed limitation and the regulation of community structure in oak savanna grassland[J].Journal of E-cology,2003,93:999-1007.
[35] 江小雷,岳静,张卫国,等.生物多样性,生态系统功能与时空尺度[J].草业学报,2010,19(1):219-229.
