青藏高原东缘典型草甸与沼泽化草甸中种子雨的差异
2012-04-12文淑均张世挺李伟杜国祯
文淑均,张世挺,李伟,2,杜国祯*
(1.兰州大学生命科学学院,甘肃 兰州730000;2.中国科学院水利部水土保持研究所,陕西 杨凌712100)
*不同的研究已证明了种子/繁殖体可利用性(seed/propagule availability)在群落结构中的作用[1-5],而对种子可利用性界定的最好方式就是对种子雨(seed rain)进行量化。种子雨是指在特定时间和特定空间从母株上散落的一定数量的种子[6],在植物生活周期(植物成体-种子扩散-种子雨-土壤种子库-幼苗-植物成体)中处于关键环节[7]。因此对于种子雨的了解,不仅可以有助于对土壤种子库的物种组成和大小的预测,进而可以对植被的更新演替趋势做出判断。
沼泽化草甸是青藏高原高寒草甸的重要组成部分,不单是优良的放牧草地,也是重要的高寒湿地,作为独特的自然生态系统,在水源涵养、水量调蓄、气候调节以及生物多样性保护等方面起了重要作用[8]。沼泽化草甸是沼泽向典型草甸的过渡类型[9],沼泽湿地退化萎缩已成为黄河源区生态恶化的重要标志之一。目前对不同高寒草甸的研究多数集中在群落结构[8,10]、牧草产量及营养动态[11]、分布范围[12]、土壤理化性质[13]、土壤微生物量、土壤酶季节动态[14,15]及不同利用程度下的碳储汇[16]。除了Ma等[17]从土壤种子库角度探讨了沼泽化草甸与典型草甸的演替关系外,在该区域有关典型草甸与沼泽化草甸种子雨的对比研究较少。因此本研究旨在了解典型草甸与沼泽化草甸中种子雨物种组成和种子雨密度差异的基础上,分析种子雨季节动态以及种子雨与地表植被的关系,并分析预测沼泽化草甸的演替趋势。
1 材料与方法
1.1 研究区自然概况
本研究在兰州大学高寒草甸与湿地生态系统定位研究站(阿孜点)进行,该站位于青藏高原东缘的甘肃省玛曲县境内,其地理坐标(N 33°39′,E 102°03′),海拔约为3 500 m;年均气温为1.2℃。年降水量约为620 mm,属高寒湿润气候;年日照时数约2 580 h,年平均霜日大于270 d。典型草甸以莎草科嵩草属(Kobresia)和禾本科的羊茅属(Festuca)、早熟禾属(Poa)、剪股颖属(Agrostis)、披碱草属(Elymuns)和菊科风毛菊属(Saussurea),以及毛茛科的银莲花属(Anemone)、毛茛属(Ranunculus)植物为优势的植被类型,并伴生其他杂类草。沼泽化草甸的指示种有藏嵩草(Kobresiatibetica)、驴蹄草属(Caltha)、垂头菊属(Cremanthodium),其他优势种包含禾本科披碱草属(Elymuns)、羊茅属(Festuca),毛茛科的银莲花属(Anemone)等,并伴随其他杂草。典型草甸与沼泽化草甸在整个生长季均有围栏保护,仅用于冬季放牧;两者被河流隔开,典型草甸地势略高于沼泽化草甸。
1.2 种子收集器的布置
2010年6月,在典型草甸与沼泽化草甸中分别选取一块植被与微地形较为均一的样地。为了避免边缘效应,样地边缘与所处草甸的边缘距离不少于30 m;每块样地中设置4条28.5 m的平行样线,样线依次编号为A,B,C,D,样线走向大致垂直于河流,其间距为5 m;每条样线上均匀放置20个15 cm×15 cm种子收集器(Astro Turf,美国生产),收集器间隔为1.5 m,每条样线上收集器依次编号为1~20。放置收集器之前用剪刀去除相同面积的地表植物,使收集器尽可能与地表齐平,用不锈钢钉将其固定于地表。此后分别于7月26日,8月12日,8月25日,9月10日,9月30日,10月15日,以收集器为单位6次收集各收集器上的种子并将其装入信封带回实验室风干,在鉴定前保持种子干燥。
1.3 植被调查
当年7月底-8月初(处于生长季高峰期,能确保所有物种鉴定)进行植被调查。在每块样地沿对角线随机放置10个50 cm×50 cm的样方框,登记样方框内所有物种,记录每物种多度、并测量其高度与盖度,盖度估计采用修正Braun-Blanquet等级法。
1.4 种子鉴定
种子鉴定时先将凋落物等杂物分离,然后选取完整的种子(种子饱满、无动物啃食痕迹),依据兰州大学生态学研究所建立的种子标本库,鉴定所有种子并计数;对属于同一属而难以区分的种子按属归类,如:禾叶嵩草(Kobresiagraminifolia),线叶嵩草(K.capillifolia),四川嵩草(K.setchwanensis);大拟鼻花马先蒿(Pedicularisrhinanthoides),甘肃马先蒿(P.kansuensis),四川马先蒿 (P.szetschuanica);钝裂银莲花(Anemoneobtusiloba),条叶银莲花(Atrullifolia);鹅绒委陵菜(Potentillaanserina),莓叶委陵菜(P.fragarioides);胡氏剪股颖(Agrostishugoniana),芒剪股颖(A.trinii);丝叶毛茛(Ranunculustanguticus),云生毛茛(R.longicaulis)等。
1.5 数据分析
在数据分析之前,参照 Kettenring和Galatowitsch[18]方法分别将每条样线上编号为1,6,11,16;2,7,12,17;3,8,13,18;4,9,14,19;5,10,15,20收集器中种子,每4个合并。
用Shannon-Wiener指数(香农指数)分别计算植被和种子雨中的多样性指数,H=-∑n
i=1p ilnpi;式中,H指多样性指数,p i指第i个物种所占比例。
物种丰富度在植被中指单个样方内的物种数,在种子雨中指收集器按4个合并后的物种数;种子雨密度换算成单位面积内种子数(粒/m2);植被中个体密度指单个样方中的植物个体数。
用P-P Plots检验数据的正态性,采用独立样本t-检验分别对典型草甸与沼泽化草甸的植被和种子雨中的物种丰富度、多样性指数和密度进行分析。所有分析均在SPSS 16.0(SPSS Inc.,Chicago,USA)中完成。
典型草甸和沼泽化草甸中种子雨与植被的物种组成相似性用无度量多维度标定排序(nonmetric multidimensional scaling,NMDS)进行验证。NMDS是一种能在多维度空间展现相似矩阵并能维持原有项目之间关系等级的非参数排序技术[19]。最终NMDS轴值表示有相似物种的点在排序空间越近,具体参考Ma等[17]。
2 结果与分析
2.1 植被概况
在2块样地中累计调查到76物种,其中共同种有38种,只在典型草甸与沼泽化草甸中出现的物种分别为22和16种。均值比较结果表明,2块样地中物种丰富度、香农指数及植株个体密度差异均不显著(表1)。NMDS结果显示,典型草甸与沼泽化草甸物种组成有明显的区别,在排序空间有明显分离(图1)。
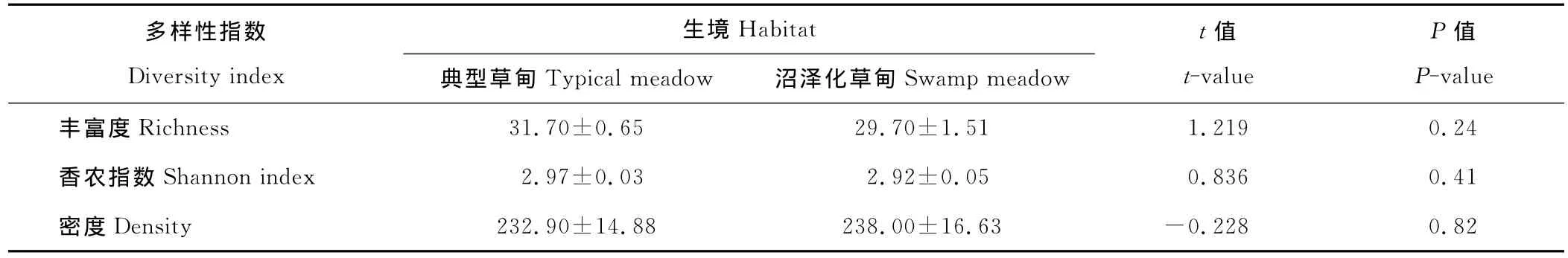
表1 典型草甸与沼泽化草甸植被的差异(平均值±标准误)Table 1 The difference of vegetation between typical and swamp meadow(Mean±SE)
2.2 种子雨概况
在沼泽化草甸中,共收集到6 751粒种子,属于14科、33种,其种子雨密度变化范围为2 288.89~5 477.78粒/m2;在典型草甸中,共收集到15 192粒种子,属于13科、30种,其种子雨密度变化范围4 944.44~10 922.22 粒/m2。具体到各科种子数量的对比,除菊科、龙胆科、蓼科和败酱科外,典型草甸中其他各科种子数均大于沼泽化草甸中的种子数(图2)。均值比较结果表明,不同草甸中收集器内物种丰富度及种子雨密度差异显著,但香农指数差异不显著(表2)。
2.3 种子雨时间动态
在6次收集时间范围内,典型草甸中种子收集器捕获的物种数由第1次的5.20种渐上升到第5次的15.25种,然后开始消退,呈单峰模式;而沼泽化草甸中种子收集器捕获的物种数由第1次的5.05种逐渐上升到第5次的11.85种(除第3次相较第2次略有下降),然后开始消退,大体呈单峰模式(图3a)。典型草甸与沼泽化草甸中种子雨密度分别由第1次的446.11和538.89粒/m2上升到第2次的1 210和1 028.9粒/m2,第3次分别下降为733.89和397.78粒/m2,由第3次开始又逐渐上升,第5次种子雨密度到达顶峰分别为3 263.9和985粒/m2,然后开始急剧下降;均呈双峰模式(图3b)。

图1 不同草甸中植被和种子雨的NMDS排序Fig.1 The NMDS ordination of vegetation and seed rain in different meadow type
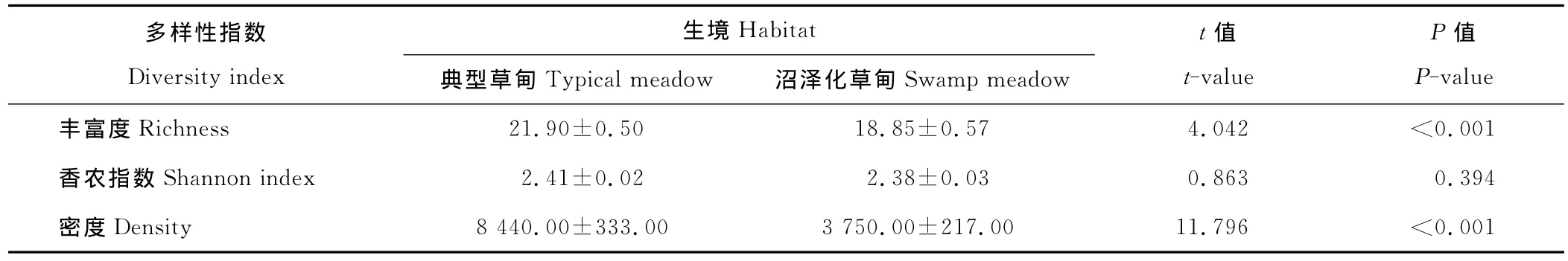
表2 典型草甸与沼泽化草甸中种子雨的差异(平均值±SE)Table 2 The difference of seed rain between typical and swamp meadow(Mean±SE)
2.4 种子雨与植被之间的关系
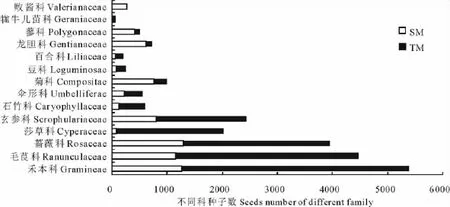
图2 各科种子数量的对比Fig.2 Comparison of seeds number in each family

图3 种子雨中物种丰富度(a)和种子雨密度(b)的时间动态Fig.3 The temporal dynamics of species richness in seed rain(a)and seed rain density(b)
3 讨论
3.1 典型草甸与沼泽化草甸中植被的差异
典型草甸与沼泽化草甸植被的物种丰富度、Shannon指数以及植株个体密度差异不显著(表1),这与Ma等[17]研究结果不一致;其可能原因是沼泽化草甸选择不一致,因沼泽化草甸是沼泽与典型草甸的过渡类型,因样地选择不一致可能会因地势增高、地下水位下降使其土壤相对干燥,因而有大量中生植物出现,所以导致本研究中沼泽化草甸植被多样性的升高。但NMDS排序显示,典型草甸与沼泽化草甸物种组成差异显著(图1),这与Ma等[17]排序结果一致;这种差异主要体现在沼泽化草甸中指示种,如藏嵩草(Kobresiatibetica)、驴蹄草 (Calthascaposa)、条叶垂头菊(Cremanthodiumlineare)。
表3 种子雨与植被的相似系数Table 3 The Srensen’s similarity coefficient between seed rain and vegetation

表3 种子雨与植被的相似系数Table 3 The Srensen’s similarity coefficient between seed rain and vegetation
项目Item St Ss Vt Vs St 1 Ss 0.806 1 Vt 0.700 0.714 1 Vs 0.535 0.667 0.688 1
3.2 典型草甸与沼泽化草甸种子雨的差异
典型草甸中收集器所收集的物种数及种子数都显著高于沼泽化草甸中的(表2),典型草甸种子雨数量的增加是由多数科属种子数的增加所引起,尤其是禾本科、毛茛科、蔷薇科种子数的增加(图2)。总结排除森林以外群落水平上种子雨的研究(表4)[18,20-38],发现仅有3处研究中种子雨密度高于本研究中典型草甸[20-22],与典型草甸种子雨密度接近的有3处[20,23-24];对比这几处高种子雨密度地区的植被,基本都会出现禾本科、菊科、豆科、蔷薇科、龙胆属植物中的1种或几种,这些植物都是种子产量较大的物种(表4)。沼泽化草甸种子雨密度与其他湿地类型对比,仅有1处比本研究高[25],有1处与之接近[18],这几处种子雨密度高的湿地中一般都有禾本科或苋科植物中的1种或几种。相较于青藏高原以外的其他高寒地区[27,29-32,34-35,38],本地区种子雨密度明显较大;尤其与加拿大极地地区相比[38],高出141~422倍。可见青藏高原在所有高寒区中种子资源极其丰富,这也是该地区高物种丰富度的基础。种子雨密度与本地土壤种子库进行对比发现,典型草甸与沼泽化草甸中种子雨的密度均高于土壤种子库的密度[17]。其可能原因有,一部分种子在形成种子库之前会被动物取食或者腐烂;其二种子库的记数采用了萌发法登记幼苗,而本地植物多为多年生植物,多为克隆繁殖方式,种子库中的种子数被低估。
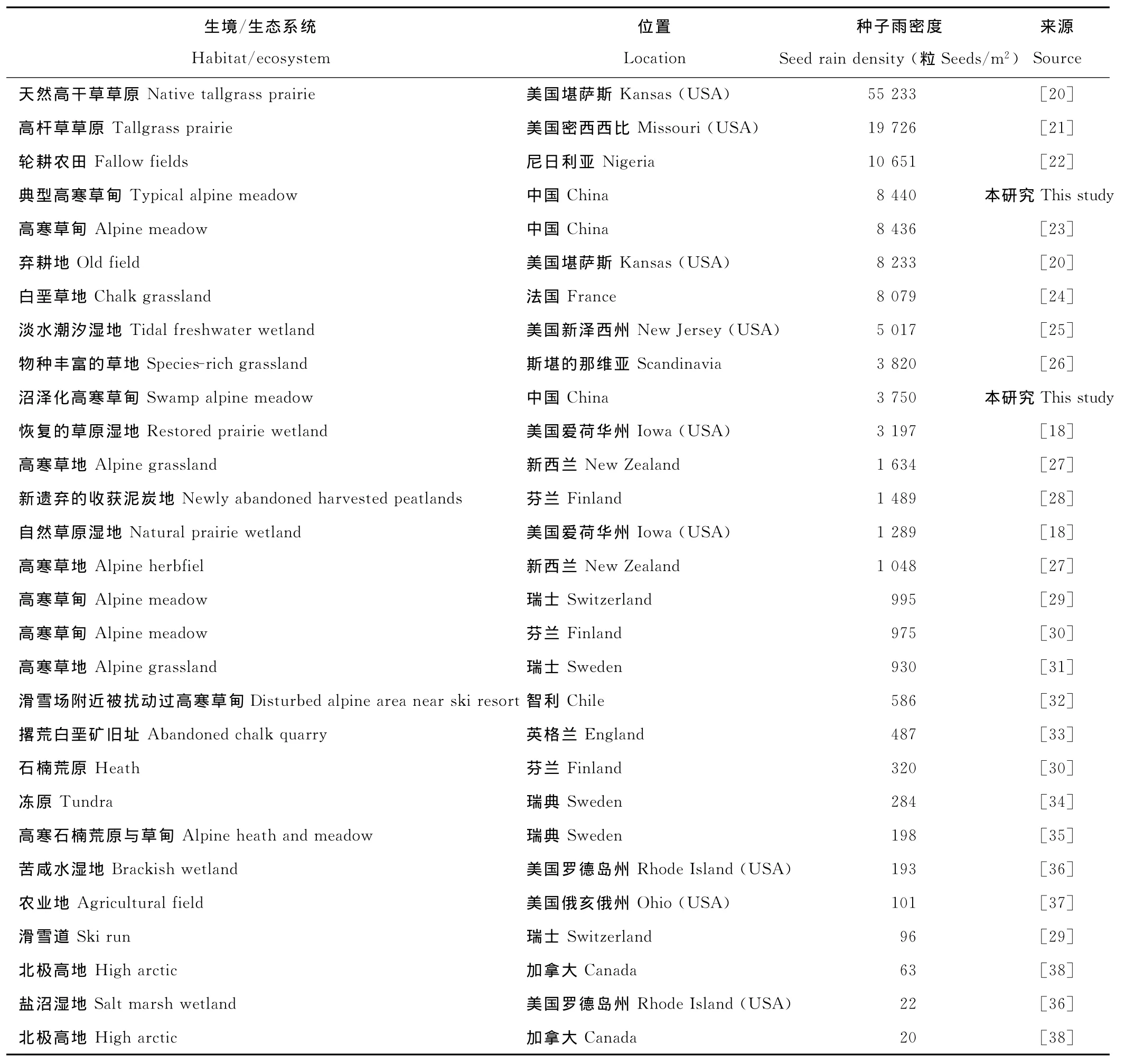
表4 不同生境中种子雨密度的对比Table 4 The comparison of seed rain in different habitat
种子雨的时间动态在不同类型的草地中呈现模式并不一致,有些研究呈单峰模式[27,29,33],有些呈双峰模式[21];本研究中典型草甸与沼泽化草甸中种子雨密度都呈双峰模式。对比单峰模式与双峰模式的研究发现,在单峰模式研究地中鉴别出种子所属物种均不超过15种;而双峰模式研究地鉴别出种子所属物种均高于30种。这种双峰模式潜在机理可能是物种花期物候在时间生态位上分化的结果,由于这些研究地物种丰富度高,生长季相对较短,而植物需要各类资源类似,时间生态位的分化使得物种共存[39]。
3.3 种子雨与植被的关系
不同研究结果表明,种子雨与植被有很强的相关性,种子雨物种的组成很大程度依赖于植被的物种组成[18,33];本研究中也是如此,典型草甸与沼泽化草甸中,种子雨与植被的相似系数分别为0.700和0.667。典型草甸与沼泽化草甸植被的相似系数为0.688,但两者种子雨的相似系数却为0.806。这种趋势在NMDS排序中得到了验证,NMDS排序结果显示典型草甸与沼泽化草甸中植被的物种组成差异显著,在排序空间有明显的分离;但种子雨的物种组成仅沿轴2略有分离,这也可以为Ma等[17]研究中种子库相似性高于植被相似性提供支持。
4 结论
典型草甸与沼泽化草甸植被中物种丰富度虽无显著差异,但物种组成上有明显的分离;典型草甸与沼泽化草甸中种子雨密度差异显著,但种子雨的相似性高于植被的相似性,若沼泽化草甸水分持续丧失演变成典型草甸的可能性较大;相较其他不同类型草地或高寒地区,该区域种子资源丰富,因而该地区采用的补播措施用来恢复退化草甸效果需要进一步量化。
[1] Zobel M,Otsus M,Liira J,etal.Is small-scale species richness limited by seed availability or microsite availability?[J].E-cology,2000,81(12):3274-3282.
[2] Foster B L.Constraints on colonization and species richness along a grassland productivity gradient:the role of propagule availability[J].Ecology Letters,2001,4(6):530-535.
[3] Foster B L,Tilman D.Seed limitation and the regulation of community structure in oak savanna grassland[J].Journal of Ecology,2003,91(6):999-1007.
[4] Foster B L,Dickson T L.Grassland diversity and productivity:the interplay of resource availability and propagule pools[J].Ecology,2004,85(6):1541-1547.
[5] Foster B L,Dickson T L,Murphy C A,etal.Propagule pools mediate community assembly and diversity-ecosystem regulation along a grassland productivity gradient[J].Journal of Ecology,2004,92(3):435-449.
[6] Harper J L.Population Biology of Plant[M].London:Academic Press,1977.
[7] Wang B,Smith T.Closing the seed dispersal loop[J].Trends in Ecology &Evolution,2002,17(8):379-386.
[8] 李英年,赵亮,徐世晓,等.祁连山海北高寒湿地植物群落结构及生态特征[J].冰川冻土,2006,28(1):76-84.
[9] 吴征镒.中国植被[M].北京:科学出版社,1980.
[10] 王启基,周兴民,沈振西,等.高寒藏嵩草沼泽化草甸植物群落结构及其利用[A].高寒草甸生态系统(4)[C].北京:科学出版社,1995:91-100.
[11] 吴海艳,马玉寿,董全民,等.黄河源区藏嵩草沼泽化草甸地上生物量及营养季节动态研究[J].草业科学,2009,26(1):8-12.
[12] 李春秀,孙海松.青海省西藏嵩草沼泽化草甸资源及保护[J].草业科学,2009,26(9):77-83.
[13] Wu G L,Liu Z H,Zhang L,etal.Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China[J].Plant and Soil,2010,332(1):331-337.
[14] 杨成德,陈秀蓉,龙瑞军,等.东祁连山高寒草地土壤微生物量及土壤酶季节动态特征[J].草业学报,2011,20(6):135-142.
[15] 王国荣,陈秀蓉,张俊忠,等.东祁连山高寒灌丛草地土壤微生物生理功能群的动态分布研究[J].草业学报,2011,20(2):31-38.
[16] 张凡,祁彪,温飞,等.不同利用程度高寒干旱草地碳储量的变化特征分析[J].草业学报,2011,20(4):11-18.
[17] Ma M J,Zhou X H,Du G Z.Soil seed bank dynamics in alpine wetland succession on the Tibetan Plateau[J].Plant and Soil,2011,346(1):19-28.
[18] Kettenring K,Galatowitsch S.Seed rain of restored and natural prairie wetlands[J].Wetlands,2011,31(2):283-294.
[19] Legendre P,Legendre L.Numerical Ecology[M].New York:Elsevier,1998.
[20] Schott G,Hamburg S.The seed rain and seed bank of an adjacent native tallgrass prairie and old field[J].Canadian Journal of Botany,1997,75(1):1-7.
[21] Rabinowitz D,Rapp J K.Seed rain in a north American tall grass prairie[J].Journal of Applied Ecology,1980,17(3):793-802.
[22] Ekeleme F,Okezie A I,Isichei A O,etal.Influence of fallow type and land-use intensity on weed seed rain in a forest/savanna transition zone[J].Weed Science,2000,48(5):604-612.
[23] 邓自发,谢晓玲,王启基,等.高寒小嵩草草甸种子库和种子雨动态分析[J].应用与环境生物学报,2003,9(1):7-10.
[24] Chabrerie O,Alard D.Comparison of three seed trap types in a chalk grassland:toward a standardized protocol[J].Plant E-cology,2005,176(1):101-112.
[25] Leck M A,Simpson R L.Tidal freshwater wetland zonation:seed and seedling dynamics[J].Aquatic Botany,1994,47(1):61-75.
[26] Jakobsson A,Eriksson O,Bruun H H.Local seed rain and seed bank in a species-rich grassland:effects of plant abundance and seed size[J].Canadian Journal of Botany,2006,84(12):1870-1881.
[27] Spence J R.Seed rain in grassland,herbfield,snowbank,and fellfield in the alpine zone,Craigieburn Range,South Island,New Zealand[J].New Zealand Journal of Botany,1990,28(4):439-450.
[28] Salonen V.Relationship between the seed rain and the establishment of vegetation in two areas abandoned after peat harvesting[J].Ecography,1987,10(3):171-174.
[29] Urbanska K M,Fattorini M.Seed rain in high-altitude restoration plots in Switzerland[J].Restoration Ecology,2000,8(1):74-79.
[30] Welling P,Laine K,Bakker J P.Regeneration by seeds in alpine meadow and heath vegetation in sub-arctic Finland[J].Journal of Vegetation Science,2002,13(2):217-226.
[31] Urbanska K M,Fattorini M.Seed rain in natural grassland and adjacent ski run in the Swiss Alps:a preliminary report[J].Restoration Ecology,1998,6(2):159-165.
[32] Chacón P,Cavieres L A.Recovery of alpine plants after a severe human disturbance in the Andes of central Chile[J].Arctic,Antarctic,and Alpine Research,2008,40(4):617-623.
[33] Jefferson R G,Usher M B.Seed rain dynamics in disused chalk quarries in the Yorkshire Wolds,England,with special reference to nature conservation[J].Biological Conservation,1989,47(2):123-136.
[34] Molau U,Larsson E L.Seed rain and seed bank along an alpine altitudinal gradient in Swedish Lapland[J].Canadian Journal of Botany,2000,78(6):728-747.
[35] Larsson E L.Two ways of trapping seeds in alpine environments,Lapland,Sweden[J].Nordic Journal of Botany,2003,23(3):333-343.
[36] Crain C M,Albertson L K,Bertness M D.Secondary succession dynamics in estuarine marshes across landscape-scale salinity gradients[J].Ecology,2008,89(10):2889-2899.
[37] Webster T M,Cardina J,White A D.Weed seed rain,soil seedbanks,and seedling recruitment in no-tillage crop rotations[J].Weed Science,2003,51(4):569-575.
[38] Grulke N E,Bliss L C.A note on winter seed rain in the High Arctic[J].Arctic,Arctic and Alpine Research,1983,15(2):261-265.
[39] 杜国祯.草本植物群落中种间时间生态位分化、群落结构组建和多样性维持重要机制[D].兰州:兰州大学,1992.
