新疆昭苏野生黄花苜蓿果实形态变异研究
2012-04-12于万里张博
于万里,张博
(新疆农业大学草业与环境科学学院 新疆草地资源与生态实验室(自治区重点)西部干旱荒漠区草地资源与生态实验室,新疆 乌鲁木齐830052)
*黄花苜蓿(Medicagofalcata)属豆科苜蓿属优良牧草,分布于我国的西北、东北、华北等地,尽管其产量和再生性逊色于紫花苜蓿,但抗逆性强,适应能力强,具有耐寒、抗旱、耐盐碱、耐风沙、抗病虫害等特性,是改良紫花苜蓿和抗性育种的重要种质资源[1-3]。我国黄花苜蓿野生种质资源丰富,尤其在新疆和内蒙古等地,伊犁河谷昭苏地区是黄花苜蓿资源较为丰富和集中的地区[4]。
目前,国内对黄花苜蓿在栽培驯化、种子特性、组织培养、抗性机理、遗传多样性等方面均有研究。王俊杰[5]、于林清等[6]、苏东等[7]对黄花苜蓿野生种群遗传多样性进行了研究,结果表明,黄花苜蓿野生种群内和种群间均存在丰富的变异,黄迎新等[8]对引进品种和锡盟野生黄花苜蓿的形态变异进行了研究。王红梅等[9]对二倍体和四倍体野生黄花苜蓿形态特征进行了比较研究,营养器官的差异比生殖器官显著。
种子是高等植物有性繁殖的主要方式之一,种子形态变异普遍存在于自然界种子植物中。研究表明,种群间、种群内甚至个体间种子形态都有很大差异[10-13]。其形态特征受遗传控制较强,受环境条件影响较小[14]。目前,国内对黄花苜蓿荚果和种子形态变异的研究未见报道,以新疆昭苏不同居群黄花苜蓿为研究对象,旨在揭示其荚果和种子表型遗传变异规律及天然群体间亲缘关系,以期为新疆黄花苜蓿遗传多样性研究及种质资源的开发利用提供理论依据。
1 材料与方法
1.1 试验材料
试验材料由新疆草地资源与生态实验室的草种质资源与育种研究室提供,2008年8月底采自新疆昭苏地区,共8个群体(表1)。
1.2 试验方法
本试验于2010年1-3月在实验室内进行。采用随机抽样法,每个群体随机选取20个单株,测定荚果和种子的11个形态指标,每个指标重复10次。
荚果形状、荚果颜色、种子颜色等指标采用数字编码法。荚果形状分3级,直线形为1、镰刀形为2、半圆形为3;荚果颜色分4级,黄色为1、黄褐色为2、褐色为3、黑褐色为4;种子颜色分4级,黄色为1、黄褐色为2、褐色为3、咖啡色为4。
直线形荚果长即荚果中线的长度,镰刀形和半圆形荚果利用细线绕着荚果弯曲,最后将细线拉直测得的长度即为荚果长度。荚果宽即荚果的最宽处。种子长、宽即种子的最长、最宽处。均用游标卡尺测量,测量精度为±0.02 mm。
因野外采样,种子量较少,种子重量采用百粒重表示,每个群体分单株取50粒种子,用万分之一电子天平称量。称量精度为0.000 1 g。
剥离荚果,记录每个荚果所含种子粒数。

表1 黄花苜蓿居群点采样信息Table 1 The sampling site information of M.falcata
1.3 数据分析
采用SPSS 17.0线性模型嵌套设计方差分析[15],研究群体间和群体内单株间2个水平上的差异。荚果和种子性状观测值的线性模型为:Y ijk=μ+Pi+T j(i)+eijk。其中,Y ijk为第i个群体第j个单株的第k个观测值,μ为总均值;Pi为第i个群体的效应值;T j(i)表示第i个群体内的第j个单株的效应值;eijk为试验误差。
用主成分分析表型差异形成的主要原因,相关性分析各性状之间以及与地理因子之间的相关关系,数据标准化后用算术平均法(UPGMA)对不同群体进行聚类分析,Excel作图。
2 结果与分析
2.1 黄花苜蓿果实性状的差异分析
试验结果显示,荚果形状多为直线型和镰刀形;荚果颜色为黄褐色和褐色;种子形状为肾形或近似肾形;颜色主要为黄色和黄褐色(图1)。
结果表明,不同群体间,昭苏地区黄花苜蓿除荚果长宽比性状无显著差异外,其他各性状均有极显著性差异。群体内单株间除荚果宽、种子颜色和种子宽有显著性差异外,其余均无显著差异(表2)。通过群体间F值比较,荚果和种子的11个形态指标中,百粒重的F值最大(25.581),其次为种子宽(23.178),种子长(18.639),荚果宽(10.674),荚果形状(5.015),其他均小于5.0。群体内F值种子宽最大(2.666),其次是种子颜色(2.032),荚果宽最小(1.969)。
昭苏地区黄花苜蓿百粒重变异系数最大(0.540),百粒重最大为151.8 mg,最小为70.6 mg,两者相差达81.2 mg,可能有二倍体黄花苜蓿。荚果形状变异系数次之(0.535)。在群体内,种子颜色变异系数最大,平均为0.554。平均每荚含种子数4.2粒(表3)。种子长、种子长宽比、种子宽等性状变异系数较低,表明种子性状具有较高的稳定性,受较高的遗传控制。

图1 不同群体黄花苜蓿荚果和种子形态差异Fig.1 Morphological diversity of pod and seed of M.falcata in different populations

表2 黄花苜蓿荚果和种子形态差异分析Table 2 Variance analysis phenotypic characteristics of pod and seed of M.falcata
2.2 相关性分析
2.2.1 荚果和种子性状间的相关性 通过Pearson相关系数分析得出(表4):荚果形状与荚果颜色之间有极显著正相关关系,表明随着荚果弯曲度的增加,颜色逐渐加深;荚果长与荚果宽和含种子数之间呈显著正相关关系,即荚果长度越长,相应的宽度也较宽,含种子数多;荚果宽与种子长、种子宽、种子长宽比和百粒重之间有显著相关关系,表明荚果越宽,则种子相应地变长、变宽、重量增加,从相关系数可以看出,对种子的宽度影响较大,所以种子长宽比变小;荚果长宽比与种子长宽比二者也有极显著正相关关系;种子颜色的深浅与每荚果含种子数之间有极显著正相关关系,即含种子粒数较多的荚果,种子颜色较深;种子的长、宽与百粒重3个性状之间有极强的关联度,呈极显著正相关关系,种子长与百粒重r=0.98,种子宽与百粒重r=0.94,表明种子长宽能反映重量的变化规律。
2.2.2 荚果和种子性状与地理因子间的相关性 除荚果长度与海拔之间有相关关系外(表4),其他各性状与地理因子之间均无相关性。荚果长与海拔的相关系数为0.71,表明随着海拔的增加,荚果呈现逐渐变长的趋势。

表3 各群体黄花苜蓿荚果和种子形态的变异Table 3 Morphological variation of pod and seed of M.falcata different populations
2.3 主成分分析
为探明影响黄花苜蓿不同群体间差异显著的主要原因,对荚果和种子的10个性状进行主成分分析。前3个主成分的累计贡献率已达到72.903%(>70%),它们可以代替原始因子所包含的主要信息(表5)。第1主成分的贡献率为35.200%,对其作用最大的性状主要为种子宽(0.949),百粒重(0.870),种子长(0.808)等,反映的是种子大小;第2主成分的贡献率为21.994%,对其作用最大的性状为荚果长/宽(0.924),含种子数(0.697),反映的是荚果含种子数;第3主成分的贡献率为15.709%,对它作用最大的性状是荚果颜色(0.593),荚果形状(0.533),反映的是荚果的形状(表5,6)。即群体间的差异主要来源于种子大小、荚果种子数和荚果形状。
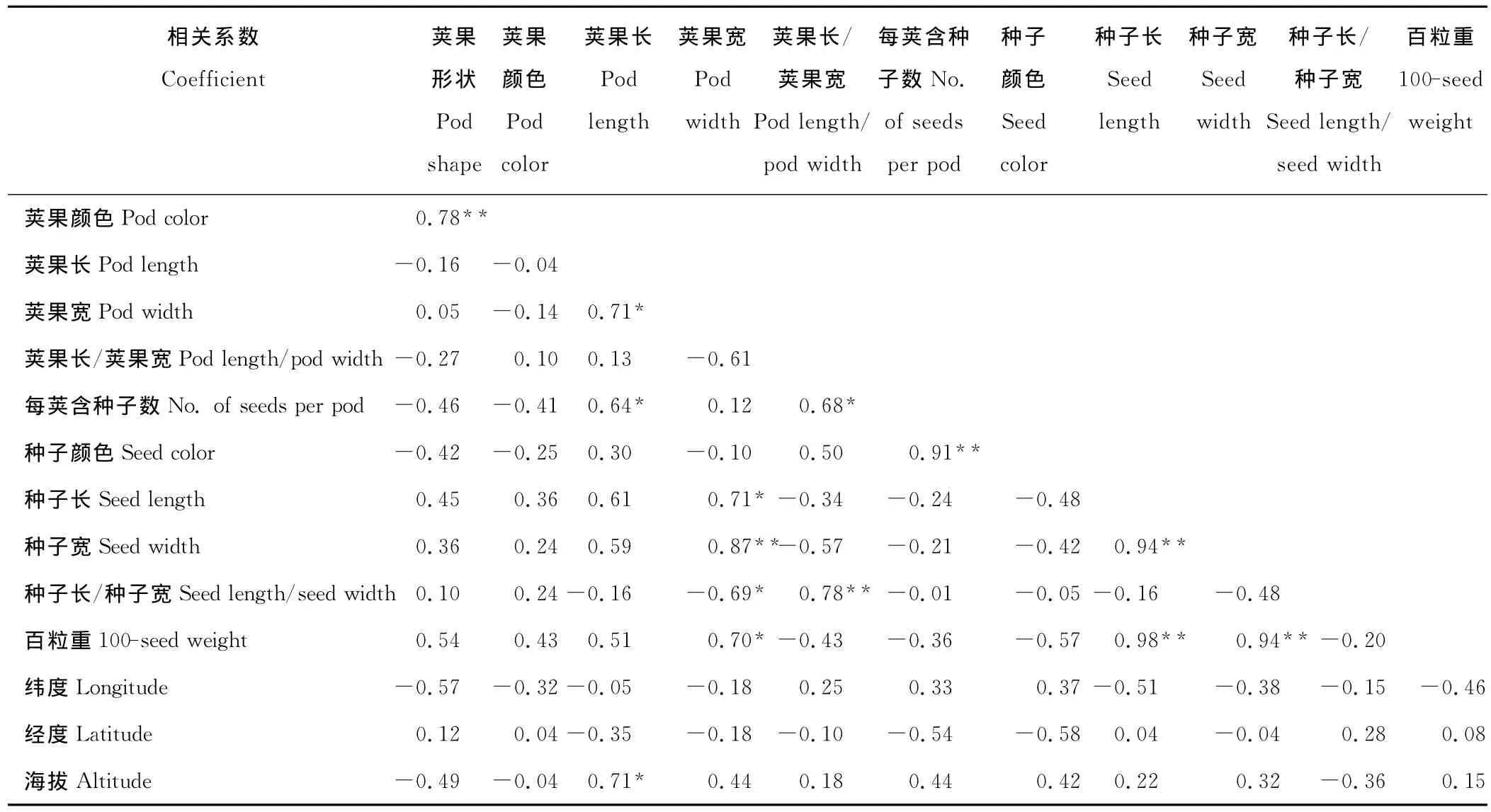
表4 黄花苜蓿荚果、种子形态及地理位置间的相关系数Table 4 Correlation coefficient among morphological trait and between morphological traits and geological factors
2.4 聚类分析
对果实10个形态指标标准化后,采用UPGMA对8个群体黄花苜蓿进行聚类分析,在欧式距离3.64处可将8个群体聚为4类(图2),其中ZMY单独为一类,该地区距其他群体较远,海拔最高,荚果大,每荚果种子粒数较多,可表述为“大荚果型”;ZMK、ZSK聚为一类,其主要特点是种子长、种子宽、百粒重数值最大,可表述为“大粒型”;ZM单独为一类,种子短小、百粒重小,可表述为“小粒型”;ZSL、ZSM、ZSJ、ZSS聚为一类,可表述为“中间型”。
3 讨论
黄花苜蓿作为野生种质资源,具有抗逆性强和适应性广的特点,国内外的研究主要集中于驯化、遗传多样性及优异特性的开发利用上。形态多样性是遗传多样性的基础,也是检测遗传变异最简便且有效的方法。但影响形态变异的因素除遗传因素外,还有生境、形态特征及观测次数的选择不同和野外调查研究中取样的局限性,一般选择比较稳定的形态指标,如叶、茎、生殖器官等[16]。

表5 黄花苜蓿果实形态相关矩阵的特征值、贡献率和累计贡献率Table 5 Eigenvalue,contribution rate and accumulative contribution rate of M.falcata
昭苏地区8个不同生境条件下的野生黄花苜蓿群体,经过长期的自然选择和适应,已产生明显的分化,果实形态特征在群体间和群体内均有丰富的遗传变异[6],表明黄花苜蓿有很强的适应性。经分析发现群体间的差异高于群体内的差异,在野生大豆(Glycinesoja)的研究中也有类似的结果[17],这与栽培品种差异主要来源于群体内的研究结果相反[18],表明野生种与栽培品种之间遗传距离较远,遗传改良的潜力巨大。因此,应尽量采集较多的野生群体,为生物多样性保护以及利用提供物质基础。
黄花苜蓿荚果各性状之间、荚果与种子性状之间以及种子各性状间都存在相关性,说明相关性状可能在基因上存在着某种连锁现象。随着荚果弯曲度的增加,荚果颜色逐渐加深;荚果长度越长,相应的宽度也较宽,含种子数增多;荚果越宽,则种子相应地变长、变宽、重量增加;荚果含种子粒数较多的,种子颜色相应较深,与百粒重无相关性,这与周刊社等[19]的研究结果相同;种子的长、宽与百粒重3个性状之间有极强的关联度,种子长宽能反映重量的变化规律,结果与何首乌(Polygonummultiflorum)的研究结果一致[20]。
地理阻隔和不同自然条件的长期作用可使果实和种子在植物居群间产生较大的差异。相关分析发现,随着海拔的增加,荚果呈现变长的趋势,其他性状与地理位置无相关性,即地理位置对黄花苜蓿果实性状的遗传变异影响较小。但通过黄花苜蓿生境地的考察以及聚类分析发现,ZMY地区海拔较高,距离人类活动区较远,受人为干扰较少,在长期的自然条件下已形成独特的大荚果型。ZSK、ZMK两地区靠近公路,人类活动频繁,又相距较近,亲缘关系较近,可能存在种质渗透。ZM地区距其他群体较远,自然条件阻碍了基因间的交流,种子百粒重小,可能有二倍体黄花苜蓿[7],需要进一步通过核型分析验证。从果实形态来看,昭苏地区黄花苜蓿的形态与地理位置有一定的关系,在麦冬(Ophiopogonjaponicus)和狗牙根(Cynodondactylon)的研究中有类似的结论[21,22]。因此,应加强此地黄花苜蓿的种质资源保护和开发利用。
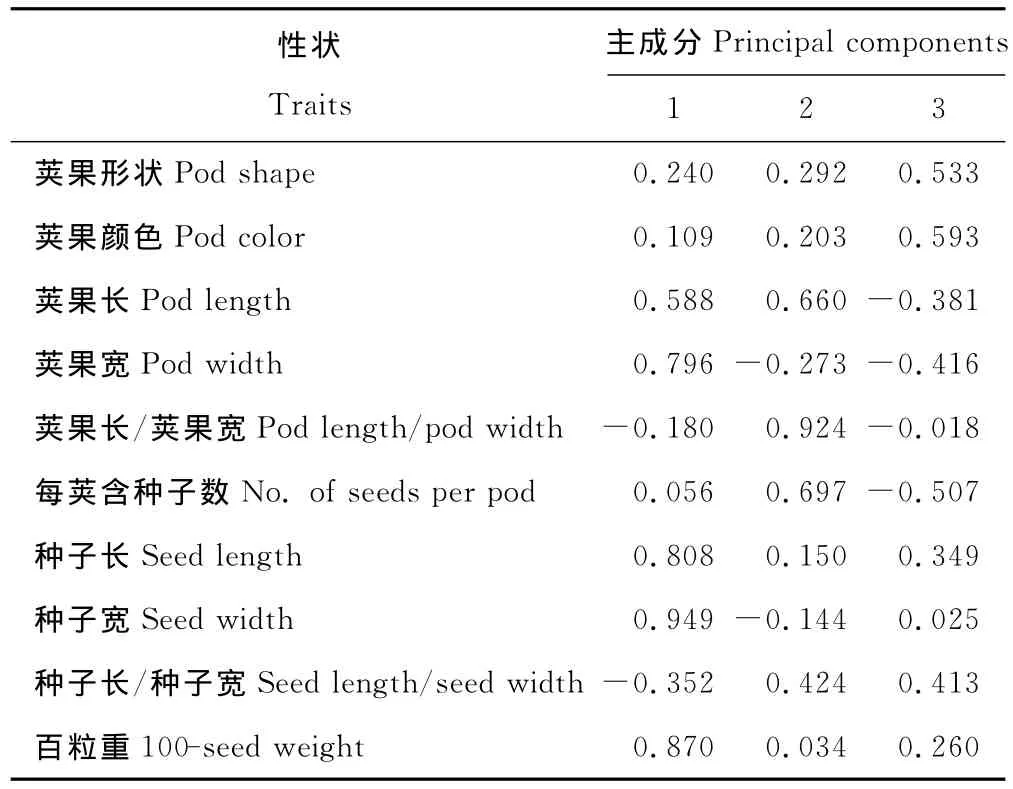
表6 黄花苜蓿果实性状的主成分因子载荷矩阵Table 6 Component matrix among populations of M.falcata

图2 基于荚果和种子形态特征的黄花苜蓿居群聚类图(UPGMA)Fig.2 The clustering dendrogram of pod and seed morphological traits of M.falcata with Euclidean distance clustered
本研究只对新疆昭苏地区野生黄花苜蓿果实形态进行了初步的研究,研究的种质资源取样范围较小,仅从荚果和种子的形态上很难完全揭示黄花苜蓿的遗传及亲缘关系。在今后的工作中应加大取样的范围,选用更多的形态指标进行研究,如叶、花等,并结合核型分析、分子标记等手段来进行,以选育优良的黄花苜蓿种质。
[1] 岳秀泉,周道玮.黄花苜蓿的优良特性与开发利用[J].吉林畜牧兽医,2004,(8):26-28.
[2] 王俊杰,云锦凤,吕世杰.黄花苜蓿种质的优良特性与利用价值[J].内蒙古农业大学学报,2008,29(1):215-219.
[3] 秦峰梅,张红香,武祎,等.盐胁迫对黄花苜蓿发芽及幼苗生长的影响[J].草业学报,2010,19(4):71-78.
[4] 贾风勤,贾娜尔,王林君.黄花苜蓿的研究进展[J].伊犁师范学院学报(自然科学版),2008,(3):22-24.
[5] 王俊杰.中国黄花苜蓿野生种质资源研究[D].呼和浩特:内蒙古农业大学,2008.
[6] 于林清,王照兰,萨仁,等.黄花苜蓿野生种群遗传多样性的初步研究[J].中国草地,2001,23(1):23-25.
[7] 苏东,周延林,于林清,等.利用SSR分析中国北方野生黄花苜蓿种群的遗传多样性[J].中国草地学报,2010,32(5):85-90.
[8] 黄迎新,周道玮,岳秀泉,等.黄花苜蓿形态变异研究[J].中国草地学报,2007,29(5):16-21.
[9] 王红梅,王俊杰,云锦凤,等.二倍体和四倍体野生黄花苜蓿形态特征比较研究[J].草地学报,2009,17(4):464-469.
[10] 孙玉玲,李庆梅,杨敬元,等.秦岭冷杉球果与种子的形态变异[J].生态学报,2005,25(1):176-181.
[11] 徐亮,包维楷,何永华.4个岷江柏松种群的球果和种子形态特征及其地理空间差异[J].应用与环境生物学报,2004,10(6):707-711.
[12] Venable D L.Size-number trade-offs and the variation of seed size with plant resource status[J].The American Naturalist,1992,140(2):287-304.
[13] Theimer T C.Intraspecific variation in seed size affects scatter-hoarding behavior of an Australian tropical rain-forest rodent[J].Journal of Tropical Ecology,2003,19(1):95-98.
[14] Baskin C C,Baskin J M.Seeds:Ecology Biogeography and Evolution of Dormancy and Germination[M].San Diego,CA:Academic Press,2001.
[15] 高忠江,施树良,李钰.SPSS方差分析在生物统计的应用[J].现代生物医学进展,2008,8(11):2116-2120.
[16] 黄春琼,周少云,刘国道,等.华南地区野生狗牙根植物学形态特征变异研究[J].草业学报,2010,19(5):210-217.
[17] 王克晶,李向华,张志卫,等.野生大豆天然群体百粒重类型组成与地理分布调查[J].大豆科学,2005,24(4):243-248.
[18] 李世雄,王彦荣,孙建华.中国苜蓿品种种子产量性状的遗传多样性[J].草业学报,2003,12(1):23-29.
[19] 周刊社,刘依兰,王彦荣.苜蓿种子产量及其构成因素的多样性研究[J].分子植物育种,2009,7(1):95-105.
[20] 施福军,王凌晖,曹福亮,等.何首乌野生种源花期与种子成熟期性状的地理变异研究[J].安徽农业科学,2009,37(25):11997-12000.
[21] 刘江,陈兴福,杨文钰,等.四川盆地麦冬种质资源的形态特征变异分析[J].草业学报,2010,19(1):143-150.
[22] 梁慧敏.不同居群狗牙根RAPD分析[J].草业学报,2010,19(1):258-262.
