Smad6信号干扰对MSCs骨向分化中Smad5及Smurf1基因表达的影响
2012-03-27刘猛董伟冯晓洁邓久鹏戚孟春李金源
刘猛 董伟 冯晓洁 邓久鹏 戚孟春 李金源
骨形态发生蛋白(bone morphogenetic proteins,BMPs)是目前刺激骨组织形成最强的生长因子,其生理效应是通过靶细胞表面Ⅰ型、Ⅱ型受体及胞内Smads信号通路来发挥的[1]。在Smads信号通路中,Smad6对BMPs信号传递起抑制作用,形成了BMPs信号传递的自身负反馈调控机制[2-4];其单独或与其它协同抑制子(如Smad泛素化调节因子-1,smad ubiguitin regulatory factor-1,Smurf1)一起,可以影响其他 Smads分子(如Smad5)的信号传递[5,6]。本研究在前期构建的小鼠 Smad6慢病毒干扰载体的基础上,探索Smad6信号干扰对小鼠间充质干细胞(MSCs)骨向分化中Smad5及Smurf1基因表达的影响,从而进一步揭示应用Smad6信号干扰促进BMP-2诱导的MSCs骨向分化的分子机制。
1 材料与方法
1.1 主要材料与试剂 携带绿色荧光蛋白(GFP)的重组干扰载体 pGCSIL-GFP-Smad6,为自行构建[7]。D/F12 小鼠骨髓MSCs专用培养基,购自广州赛业;胎牛血清购自hyclone公司;Percoll分离液购自美国PHAMACIA公司;人重组BMP-2购自北京博奥森生物技术有限公司。TrizolTMRNA Isolation Reagent,美国Gibco BRL 公司;iQ SYBR Green supermix,美国 Bio-Rad公司;上下游引物,上海生工合成。兔抗鼠单克隆抗体(一抗),购自美国SANTA CRUZ公司。
1.2 重组干扰载体对间充质干细胞(MSCs)的转染及骨向分化 密度梯度离心法培养小鼠骨髓MSCs[8],应用含15%胎牛血清的DMEM/F12培养基进行培养。取第三代MSCs用于本实验。实验分为A、B、C 3组:A组,细胞作为对照组;B组,空白载体转染+BMP-2诱导;C组,重组干扰载体转染+BMP-2诱导。转染病毒液均为50 μl/孔。转染前更换培养基,换用无血清的培养基,使细胞同步化。应用病毒载体转染B、C 2组MSCs,转染第72小时后,更换新鲜培养基,荧光显微镜下观察;并用200 ng/ml的rhBMP-2对小鼠骨髓MSCs进行骨向诱导。于BMP-2诱导第6、24小时两个时间点收获细胞,进行相关检测。
1.3 SyberGreen实时荧光定量PCR检测Smurf1的基因表达水平 参照NCBI Genebank中小鼠Smurf1和GAPDH基因序列,根据SyberGreen Real-time PCR的要求,设计待检测基因上下游引物。见表1。

表1 小鼠基因Smurf1和GAPDH的Real-time RT-PCR引物
Trizol提取总RNA,用RT-PCR kit在Rotor-Gene 3000荧光定量PCR仪上逆转录成cDNA后,取逆转录产物进行实时荧光定量RT-PCR;引物标记采用荧光染料SYBR Green。PCR扩增反应条件为:94℃ 2 min;53℃ 20 s,60℃40 s,共 45个循环。GAPDH的cDNA经倍比稀释后进行同期PCR,用于回归分析。反应结束后在PCR仪上自动读取阈值Ct;PCR产物同时进行琼脂糖凝胶电泳分析。每组细胞检测三个样本(n=3)。
1.4 Western-blot检测磷酸化 Smad5(pSmad5)、Smad5 和Smurf1蛋白水平 MSCs经病毒转染及BMP-2骨向诱导后,于两个时间点收获B、C两组细胞,提取总蛋白。测定蛋白浓度,以每孔90 pg上样,浓缩胶电泳条件为80 V、30 min;分离胶条件为150 V、2 h;恒流电转膜2.5 h。5% 牛血清白蛋白室温封闭1 h,一抗4℃过夜,二抗室温60 min,BCIP/NBT显色1 min。硝酸纤维素膜上蛋白表达的条带用自动图像分析系统(Image J)进行分析。
1.5 统计学分析应用SPSS 13.0统计软件,计量资料以表示,3组细胞基因相对浓度进行单因素方差分析,P<0.05为差异有统计学意义。
2 结果
2.1 病毒载体转染小鼠骨髓 MSCs后GFP观察 B、C 2组MSCs分别经空白载体和重组病毒载体转染,24 h后绿色荧光蛋白(GFP)开始表达,在荧光显微镜下细胞胞浆呈绿色;转染后72 h,GFP表现为强表达(图1),病毒对细胞的转染效率接近100%;在BMP-2处理后,两组细胞内GFP仍持续强表达,提示病毒载体的转染为稳定转染。

图1 病毒转染MSCs 72 h后GFP表达(荧光显微镜×100)A:B组(空白病毒载体);B:C组(重组病毒载体)
2.2 实时定量PCR检测Smurf 1基因表达 将待测样品在定量PCR仪上进行扩增反应,采集每一反应管中荧光强度的变化,并绘制动力学曲线,得出样品荧光强度增加到阈值时的Ct值,并由分析软件自动计算出与内参管家基因GAPDH比较的ΔCt值,以及与A组(无病毒转染及BMP-2处理)ΔCt值比较的ΔΔCt。样本相对浓度 =1/2ΔΔCt。
在两个时间点,3组Smurf 1相对浓度经单因素方差分析均有统计学意义(P<0.01);除BMP-2诱导6 h时A、B 2组间差异无统计学意义外,两个时间点各组两两比较差异均有统计学意义(P<0.01)。提示 BMP-2诱导显著提高了 MSCs中Smurf1基因的表达,而Smad6 RNA干扰使其表达进一步显著上升;随着时间的延长,这种变化更明显。见表2,图2。
表2 SyberGreen实时定量PCR检测Smurf 1基因表达n=3,±s

表2 SyberGreen实时定量PCR检测Smurf 1基因表达n=3,±s
注:与 A 组比较,*P <0.01;与 B 组比较,#P <0.01
?
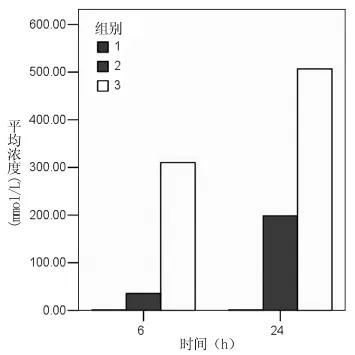
图2 实时定量PCR检测Smurf 1基因相对浓度
2.3 Western-blot检测pSmad5、Smad5和Smurf1蛋白水平 在BMP-2处理后两个时间点,B组(空白载体)、C组(重组载体)细胞Smad5总蛋白水平变化不显著,未受Smad6信号干扰的影响。而pSmad5的蛋白水平在C组比B组则明显提高,蛋白条带灰度值在两个时间点分别上升了26.31%和48.21%。上述结果说明,Smad6信号干扰虽然对Smad5总蛋白水平没有影响,但明显提高了磷酸化Smad5的水平,即促进了Smad5磷酸化,从而增强BMPs的信号传导。Smurf1蛋白水平的变化规律与pSmad5相似。C组Smurf1蛋白水平在两个时间点比B组均明显提高,蛋白条带灰度值分别上升了63.72%和58.65%。提示Smad6干扰后,Smurf1蛋白代偿性表达增高,在一定程度上补偿了因Smad6干扰对BMPs信号负调控的抑制。见图3。

图3 Western blot检测pSmad5、Smad5、Smurf1的蛋白水平
3 讨论
Smad6是BMPs信号通路中重要的自身负反馈调控因子[2-4,9]。在 BMPs刺激下,胞内 Smad1、Smad5 等受体型 Smad(R-Smad)与Smad4结合成异源多聚体,易位进入核内,作用到靶基因特异序列,发挥BMPs各种生理效应。Smad6基因上有SBE序列(smad binding element),能够与R-Smad直接结合;并激活了 Smad6基因转录,使其表达水平迅速升高[3,9],发挥对BMPs信号的负调节作用。
研究表明,Smad6可在胞浆内直接与BMPs I型受体结合,阻碍R-Smad(Smad1、Smad5)磷酸化(激活);也可与 Smad1、Smad5直接结合,阻碍它们与Smad4形成异源多聚体,进而发挥阻碍 BMPs 信号传递的作用[2,3,9]。
同时,Smad6的表达和活性受许多其它信号分子的协同调节;在这些协同分子中,大多数是Smad6的协同抑制子(corepressor);Smurf1便是其中之一。Smurf1高表达在体外可抑制成骨细胞前体细胞2T3骨向分化;在体内会使成骨细胞增殖和分化显著下降,并导致骨小梁体积及骨形成速度显著降低[10]。而Smurf1突变失活则可增强BMP-2诱导的成骨细胞前体C2C12和2T3细胞的骨向分化[11]。Smurf1可直接与BMPs I型受体及Smad1、Smad5结合,使蛋白泛素化(uniquitiation),从而导致蛋白降解,阻碍BMPs信号传导。Smurf1还可与Smad6形成复合体,使其与I型受体及Smad1、Smad5的结合更为容易;这样,一方面发挥Smad6阻碍BMPs信号传导的作用,另一方面发挥Smurf1对蛋白的降解作用,使对BMPs信号的负调控更强[5,6,10-12]。在共同调控中,Smurf1 与 Smad6 复合体对磷酸化的Smads的作用比非磷酸化的Smads作用更强。
在前期研究中,我们已证实Smad6信号干扰可有效促进BMP-2诱导的MSCs骨向分化中(另文报道);然而,Smad6信号干扰会对Smads信号通路中其它信号分子(如Smad5、Smurf1)产生怎样的影响,正是本研究所要解决的问题。
本研究结果表明,在BMP-2诱导的MSCs骨向分化中,Smad6信号干扰可有效促进Smad5蛋白磷酸化,而对Smad5总蛋白量无影响。提示Smad6信号干扰有效解除了其对Smad5磷酸化的抑制,从而促进了信号传递,增强了BMPs的效应,表现为MSCs骨向分化增加。同时,本研究表明,Smad6信号干扰一定程度上增加了Smurf1的mRNA及蛋白表达。由于Smurf1是Smad6的协同抑制子,发挥着对BMPs信号传递的抑制作用,因而上述结果提示机体内部存在一定的代偿机制,在Smad6被干扰后,Smurf1蛋白代偿性表达增高,在一定程度上补偿了Smad6分子对BMPs信号的负调控。
鉴于BMPs体内信号转导的复杂性,Smad6引号干扰不仅能促进BMP-2诱导的骨向分化,影响Smad5磷酸化及Smurf1基因表达,还可能对下游一系列信号分子及效应基因产生影响,从而发挥BMPs促进骨再生的效应。然而有哪些信号分子及效应基因会受到影响?它们的基因表达会发生怎样的变化?尚待进一步深入研究。
1 Miyazono K,Maeda S,Imamura T.BMP receptor signaling:transcriptional targets,regulation of signals,and signaling cross-talk.Cytokine Growth Factor Rev,2005,16:251-263.
2 Ishida W,Hamamoto T,Kusanagi K,et al.Smad6 is a Smad1/5-induced smad inhibitor.Characterization of bone morphogenetic protein-responsive element in the mouse Smad6 promoter.J Biol Chem,2000,275:6075-6079.
3 Goto K,Kamiya Y,Imamura T,et al.Selective inhibitory effects of Smad6 on bone morphogenetic protein type I receptors.J Biol Chem,2007,282:20603-20611.
4 Bai S,Shi X,Yang X,et al.Smad6 as a transcriptional corepressor.J Biol Chem,2000,275:8267-8270.
5 Sangadala S,Metpally RP,Reddy BV.Molecular interaction between Smurf1 WW2 domain and PPXY motifs of Smad1,Smad5,and Smad6-modeling and analysis.J Biomol Struct Dyn,2007,25:11-23.
6 Murakami G,Watabe T,Takaoka K,et al.Cooperative inhibition of bone morphogenetic protein signaling by Smurf1 and inhibitory Smads.Mol Biol Cell,2003,14:2809-2817.
7 于静,戚孟春,邓久鹏,等.小鼠Smad6慢病毒干扰载体的构建及筛选.生物医学工程学杂志,2010,27:1100-1104.
8 韩立赤,胡静,戚孟春,等.密度梯度离心和贴壁法分离大鼠骨髓间充质干细胞增殖活性和成骨功能比较.现代口腔医学杂志,2005,19:287-290.
9 Wang Q,Wei X,Zhu T,et al.Bone morphogenetic protein 2 activates Smad6 gene transcription through bone-specific transcription factor Runx2.J Biol Chem,2007,282:10742-10748.
10 Zhao M,Qiao M,Harris SE,et al.Smurf1 inhibits osteoblast differentiation and bone formation in vitro and in vivo.J Biol Chem,2004,279:12854-12859.
11 Zhao M,Qiao M,Oyajobi BO,et al.E3 ubiquitin ligase Smurf1 mediates core-binding factor alpha1/Runx2 degradation and plays a specific role in osteoblast differentiation.J Biol Chem,2003,278:27939-27944.
12 Shen R,Chen M,Wang YJ,et al.Smad6 interacts with Runx2 and mediates Smad ubiquitin regulatory factor 1-induced Runx2 degradation.J Biol Chem,2006,281:3569-3576.
