外源多胺对薄皮甜瓜花芽分化及花发育的影响
2012-02-23
(沈阳农业大学园艺学院,设施园艺省部共建教育部重点实验室,辽宁省设施园艺重点实验室,辽宁沈阳110866)
多胺是生物体代谢过程中产生的具有调控作用的低分子量脂肪族含氮碱,包括腐胺(Put)、亚精胺(Spd)、精胺(Spm)和尸胺(Cad)等。它们广泛存在于各种植物中,与植物的生长发育密切相关。植物的性别分化与激素和多胺有关,已经在很多植物上得到证明。外源多胺能显著促进红富士苹果花芽分化(Altamura & Tomassi,1998),其中Put 被认为是形成花的主要多胺。也有人认为,Put 对板栗结实花分化有显著抑制作用,而Spd 对结实花分化有促进作用(张新生等,2005)。在油菜中发现较低含量的多胺(主要是Put 和Spd)有利于油菜花芽分化的起始,含量的增加有利于花芽的发育,抽薹越早的品种其体内多胺含量越早达到较高水平(艾育芳 等,2011)。开花时期,在花的萼片、花瓣和性器官中,多胺水平上升,子房中积累大量的Spm(de Dios et al.,2006)。高含量的Cad 有利于甜瓜雄蕊的成熟,高含量的Spd 可能有利于早期雌蕊的发育(张建农和李计红,2007)。在盛花期之前喷施Spm 可以有效地延缓花的衰老(Seo et al.,2009)。目前,外源多胺对甜瓜花芽分化进程和时期的影响还不明确,本试验通过外源喷施多胺,研究甜瓜叶片和花芽中内源多胺和激素水平的变化,以期从生理水平上揭示外源多胺对内源多胺和植物激素及甜瓜花芽分化的影响,也为甜瓜性别分化外源调控提供依据。
1 材料与方法
1.1 试验材料
供试材料为薄皮甜瓜(Cucumis meloL.var.makuwaMakino)永甜3 号,来自黑龙江省齐齐哈尔市永和甜瓜经济作物研究所。
1.2 试验方法
2008年8月播种,穴盘基质育苗,每盘50 株。8月14日子叶展开时开始喷施外源多胺(Sigma公司),设4 个处理:T1,Put 1×10-3mol·L-1;T2,Put 1×10-4mol·L-1;T3,Spd 1×10-3mol·L-1,T4,Spd 1×10-4mol·L-1。以喷清水为对照(CK)。每处理3 盘,3 次重复。以后每7 d 喷施1次,共喷3 次。子叶展开当天开始取样,每隔5 d 取1 次样。当甜瓜幼苗4 片真叶时,于9月4日定植于高效节能日光温室,随机区组排列,每5 畦为1 个小区,3 次重复,高畦双行栽培,株距40 cm,黑色地膜覆盖,滴灌,三蔓整枝,爬地栽培,田间管理与生产相同。
1.3 调查项目
1.3.1 花芽分化状态和时期的显微结构观察、结实花花期及第1 节位结实花率调查 子叶展开时开始取样(张多娇 等,2009),每隔5 d 取1 次样,常规石蜡切片法观察花芽分化状态与时期。结实花开花日期和节位:结实花开花当天采用防落素喷花,并挂标签记录开花日期及节位,统计第1 结实花开花日期、初花期(30%结实花开放)和盛花期(70%结实花开放)。
1.3.2 内源激素和内源多胺的测定 子叶展开时开始取样,每隔5 d 取1 次样,取花芽和幼叶1.0 g,液氮速冻后贮藏在-80 ℃冰柜中。GA3、IAA、ZT 和ABA 等内源激素含量测定采用高效液相色谱法(张建农和李计红,2007)。色谱条件:色谱柱Spherisorb C18,柱温为25 ℃,洗脱液为甲醇∶水=64V∶36V,流速0.5 mL·min-1,紫外检测器,检测波长230 nm,进样量20 μL,外标法定量。提取方法:称取植物材料0.5 g,加入2 mL 预冷的5% HClO4研磨,研磨混合液在冰浴中放置1 h 后离心(0~4 ℃,48 000×g,20 min)。取上清液1 mL,加入2 mol·L-1的NaOH 2 mL 和苯甲酰氯20 μL,涡漩20 s 后30 ℃下保温60 min;加入饱和NaCl 2 mL 和乙醚2 mL 混合萃取苯甲酸多胺,振荡后1 500×g 离心5 min。收集1 mL 醚相,热气流(50 ℃)蒸干,-20 ℃保存。高效液相色谱测定前用100 μL 甲醇溶解样品,取20 μL 进样。
用高效液相色谱法(HPLC)测定Spm、Spd 和Put 含量(郝建军和刘廷吉,2001)。
试验数据采用Excel、DPS 软件进行相关分析,Duncan’s 多重差异显著性分析。
2 结果与分析
2.1 外源多胺对薄皮甜瓜花芽分化进程的影响
外源多胺对薄皮甜瓜花芽分化进程的影响见表1。8月14日未进行多胺处理之前,各处理均为子叶展开,石蜡切片观察,花芽未分化。经过5 d,8月19日取样时,即喷施1 次多胺后,各处理仍相同,均处于花芽分化初期。8月24日后,各处理花芽分化进程产生差距,表现为T1和T4 处理均进入花冠裂片分化期;T3 处理次之,进入萼片分化期;而T2 处理和CK 仍停留在花芽分化初期。到9月3日时,仍是T1 处理花芽分化最快,已经发育形成结实花;T4 处理其次,进入雌蕊分化期;然后是T3 处理,CK 和T2 处理花芽分化最慢。
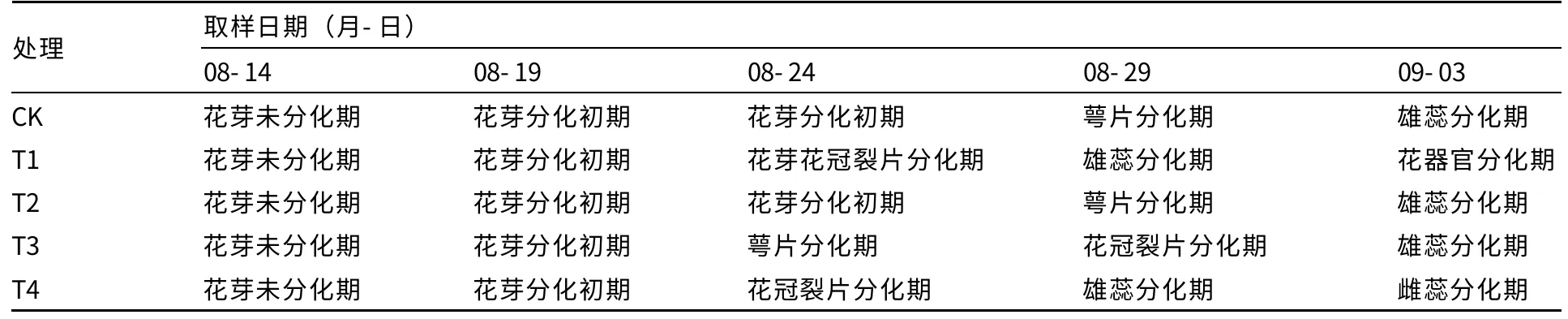
表1 外源多胺对薄皮甜瓜花芽分化进程的影响
2.2 外源多胺对薄皮甜瓜结实花花期和子蔓第1 节位结实花率的影响
由表2 可以看出,经过外源多胺处理后,T1 和T4 处理的花期基本相同或相近,第1 结实花开花期比CK 和其他两个处理均早1~2 d;T2 处理花期最晚,盛花期比T1 处理晚6 d。CK与T3 处理第1 结实花开花期、初花期、盛花期均相同。T4 处理的子蔓第1 节位结实花率最高,T1 处理次之,T3 处理最低,仅为66.67%。
由此可以看出,经过不同浓度多胺处理后,T1 和T4 处理的花期最早,子蔓第1 节位结实花率较高;CK 与T3 处理花期相同,子蔓第1 节位结实花率较低;T2 处理花期虽然最晚,但是子蔓第1 节位结实花率极显著高于CK 和T3 处理。
2.3 外源多胺对薄皮甜瓜叶片中内源激素含量的影响
2.3.1 对GA3含量的影响 从图1-a 可以看出,子叶展开未进行多胺处理时,叶片中GA3含量较低,进入花芽分化期后迅速升高,各处理在苗龄15~20 d 时均达到峰值,其中T4 处理叶片GA3含量在苗龄15 d 时(即雄蕊分化期)达到峰值,为195.1 μg·g-1(FW),显著高于其他处理,但此后下降;其他处理在苗龄20 d 时达到峰值,CK 和T1 处理GA3含量低于其他处理。说明外源喷施多胺后,GA3含量产生变化,其中喷施1×10-4mol·L-1的Spd(即T4 处理)后,GA3含量明显高于其他处理,较早达到峰值,同时T4 处理花芽分化速度快于其他处理。因此,外源多胺通过影响GA3含量影响了花芽分化,外源喷施多胺后GA3含量增高,花芽分化加快。
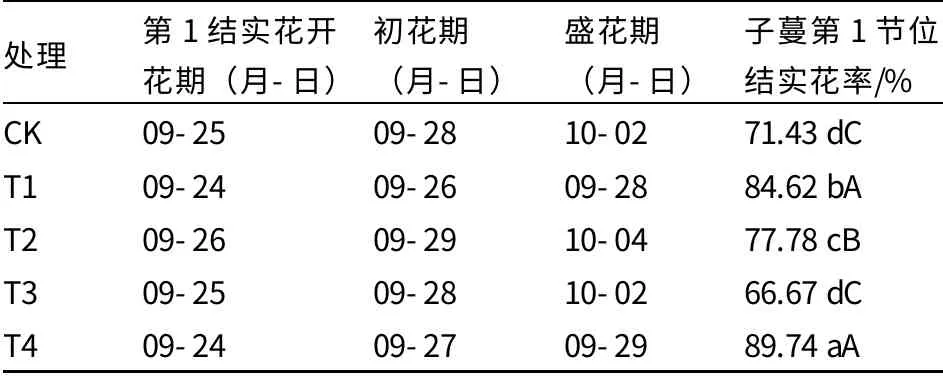
表2 外源多胺对薄皮甜瓜结实花花期和子蔓第1 节位结实花率的影响
2.3.2 对IAA 含量的影响 由图1-b 可知,子叶展开时叶片中IAA 含量处于较高水平,为8.817 μg·g-1(FW);在苗龄5 d 时,各处理均进入花芽分化初期,叶片中IAA 含量急剧降低;苗龄10 d 时,各处理IAA 含量均有所上升,升高幅度不同,T1 和T4 处理显著高于其他处理,此时正值第1 片真叶展开,植株生长迅速,石蜡切片显示T1 和T4 处理正进入花冠裂片分化期,快于其他处理;到苗龄20 d 时,各处理IAA 含量均有所下降。因此,外源喷施多胺后对IAA 含量变化影响不大,IAA 含量变化可能与植株生长速度有关。
2.3.3 对ZT 含量的影响 由图1-c 可知,叶片中ZT 含量在整个花芽分化期呈下降的趋势,子叶展开时ZT 含量处于较高水平,为26.178 μg·g-1(FW),苗龄5 d 时进入花芽分化初期,各处理ZT 含量变化幅度较小,苗龄10~15 d 时,各处理ZT 含量均有所下降,其中T1 和T4 处理下降幅度较大,在苗龄15 d 时T1 和T4 处理ZT 含量到达最低值,苗龄20 d 时ZT 含量有所上升,而其他3 个处理均在苗龄20 d 时到达最低值,这可能与多胺处理后花芽分化速度和进程有关。T1 和T4 处理较其他3 个处理提前进入雄蕊分化期。因此,外源多胺处理影响叶片中ZT 含量,外源喷施1×10-3mol·L-1的Put(T1)和1×10-4mol·L-1的Spd(T4)使叶片中ZT 含量降低,花芽分化速度快于其他处理。
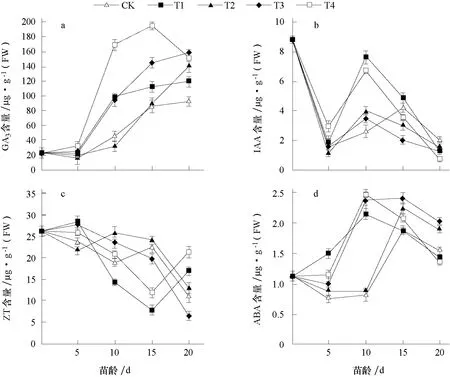
图1 外源多胺对薄皮甜瓜叶片中内源激素含量的影响
2.3.4 对ABA 含量的影响 由图1-d 可知,各处理叶片中ABA 含量变化幅度较小,子叶展开时ABA 含量处于相对较低水平,苗龄5~15 d 时各处理ABA 含量均有所上升,其中T1 和T4处理在苗龄10 d 时到达峰值,此后下降;而其他3 个处理均在苗龄15 d 时到达峰值,此后下降。因此叶片中ABA 含量变化与外源喷施多胺后花芽分化进程有关,T1 和T4 处理花芽分化速度较快。
2.4 外源多胺对薄皮甜瓜花芽中内源多胺含量的影响
由图2-a 可知,子叶展开当天,即花芽分化的生理分化期时,花芽中内源Put 含量处于相对较高水平,为736.324 nmol·g-1(FW),说明花芽分化开始前需要积累大量的内源Put。此后花芽分化开始,各处理内源Put 含量迅速下降,在花芽分化初期(苗龄5 d 和10 d)各处理的内源Put 含量下降到最低值,其中CK 在苗龄10 d 时内源Put 含量仅为14.087 nmol·g-1(FW),苗龄10~15 d 时各处理内源Put 含量均呈上升趋势,苗龄15~20 d 期间CK、T1 和T3 处理内源Put含量均保持上升趋势,而T2 和T4 处理有下降趋势。这可能与多胺处理后花芽分化进程不同有关,结合石蜡切片的花芽分化进程发现,雄蕊分化期时内源多胺的含量普遍较低,花芽分化后进入花器官分化期时内源多胺的含量相对较高。
图2-b 中,在花芽分化的生理分化期时,内源Spd 含量处于较低水平,花芽分化初期各处理内源Spd 含量变化幅度较小,进入花芽形态分化期后内源Spd 含量有不同幅度上升,苗龄10 d时T1 和T4 处理内源Spd 含量达到峰值,而其他3 个处理在苗龄15 d 时才达到峰值,这可能与外源喷施多胺后花芽分化速度有关,T1 和T4 处理花芽分化速度快于其他处理,但在两性分化期时T1 和T4 处理内源Spd 含量均有所下降,苗龄20 d 时T1 和T4 处理的内源Spd 含量高于其他处理,这可能与花芽分化进程有关,此时T1 和T4 处理完成花芽分化,已进入花器官分化期。因此,外源喷施多胺后影响内源Spd 含量变化,T1 和T4 处理的内源Spd 含量较早达到高峰。
由图2-c 可知,花芽中内源Spm 含量变化呈下降趋势,在花芽分化的生理分化期时内源Spm 含量处于较高水平,进入花芽形态分化期后内源Spm 含量有下降趋势,苗龄5~15 d 时内源Spm 含量变化不规律,但T2 处理的内源Spm 含量高于其他处理。外源多胺对内源Spm 含量影响不大。
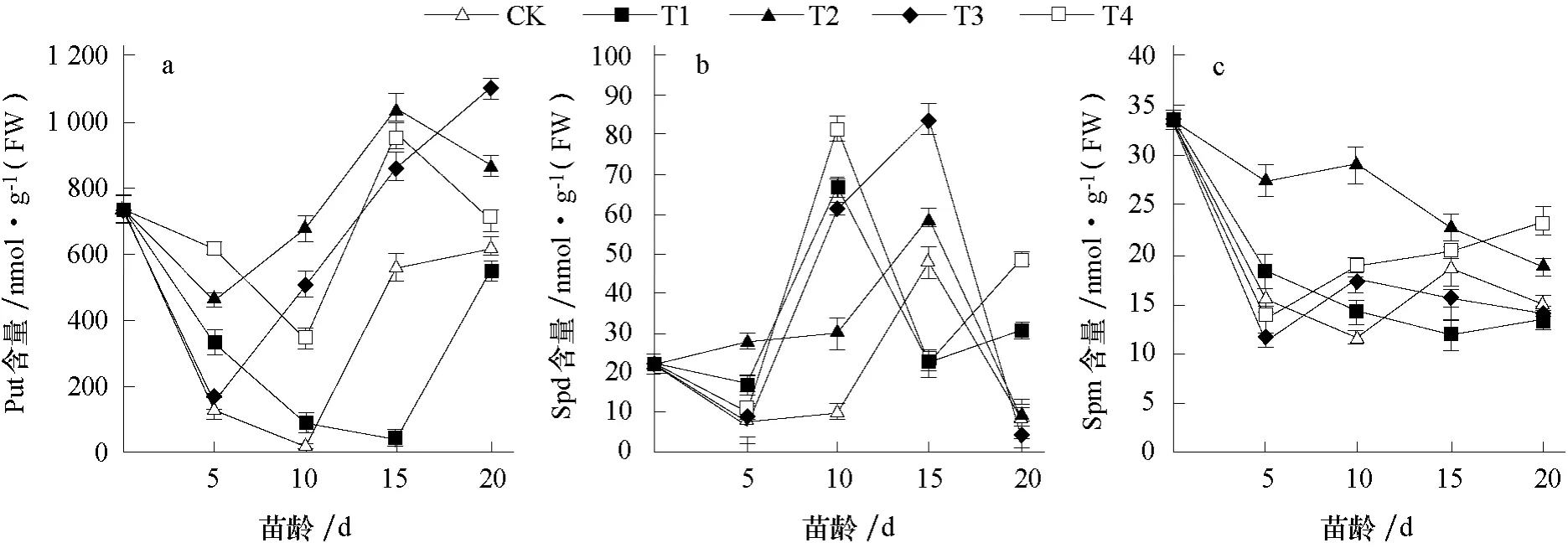
图2 外源多胺对薄皮甜瓜花芽中内源多胺含量的影响
叶片中3 种内源多胺的变化趋势与花芽中的大致相同,但花芽中3 种内源多胺的含量远高于叶片中,其中花芽中内源Put 含量约为幼叶中的100 倍,叶片中的多胺可能大量运往花芽中以供给花芽分化。并且,外源喷施不同多胺后,内源多胺含量在大部分时期高于对照,可能是喷施外源多胺增加了内源多胺的含量。
3 结论与讨论
3.1 外源多胺对薄皮甜瓜结实花花期、子蔓第1 节位结实花率和花芽分化进程的影响
不同浓度、不同种类的外源多胺对薄皮甜瓜花芽分化影响不同。外源喷施Put 和Spd 处理均能显著增加核桃的结实花数量,提高雌雄花比例(徐继忠 等,2004)。本试验结果表明,外源喷施高浓度Put(1×10-3mol·L-1)和低浓度Spd(1×10-4mol·L-1)有利于提高甜瓜子蔓第1 节位结实花率,促进花芽分化,使结实花花期提前。
3.2 外源多胺对薄皮甜瓜叶片中内源激素含量的影响
植物激素与花芽分化关系密切。GA3与第1 结实花的发育有关(汪俏梅和曾广文,1997);IAA 可能是黄瓜性别发育的关键性激素(陈学好 等,2002),外源喷施Spd 和Spm 可提高内源IAA 含量(桂仁意 等,2003);ZT 有助于甜瓜的性器官分化及发育(李计红,2006);不同内源ABA 含量在花芽分化不同时期对苹果梨的影响不同(李秉真 等,2000);在花芽分化期茎尖IAA、GA 含量保持较低的水平和适宜的PAs 含量有利于花芽分化(王廷芹和杨暹,2008)。通过本试验可得出,外源喷施多胺使叶片中GA3含量增高,花芽分化加快。喷施高浓度Put(1×10-3mol·L-1)和低浓度Spd(1×10-4mol·L-1)后,叶片中IAA 含量高于其他处理,ZT 含量降低,ABA 含量提前达到峰值,这与两处理的花芽分化速度快于其他处理有关。
3.3 外源多胺对薄皮甜瓜花芽中内源多胺含量的影响
外源喷施一定浓度的Put 和Spd 后,内源多胺的含量也随之受到影响,从而影响花的性别分化和花发育(汪俏梅和曾广文,1997;徐继忠 等,2004)。前人对荔枝花芽分化研究时发现花芽和叶片的Spd 和Spm 比值在1~3 范围,说明成熟组织也合成多胺,而且叶片的数量远远超过顶芽或花芽数量,它们合成的多胺总量比花芽多(肖华山 等,2006),因此,叶片多胺的合成对花芽的分化起着至关重要的作用。本试验中,外源喷施多胺后,花芽中内源多胺含量在大部分时期高于对照,说明外源多胺施用促进了内源多胺的合成。
外源多胺可能通过影响内源激素含量进而调控基因表达影响花芽分化,或通过内源多胺含量进而影响激素平衡而调控基因表达影响花芽分化,或内源多胺含量升高直接调控基因表达影响花芽分化,这些还需要进一步试验予以确定。
艾育芳,陈观水,周以飞,潘大仁,宁文君.2011.油菜早熟突变体成花过程中叶片多胺代谢研究.中国农学通报,27(12):101-105.
陈学好,曾广文,曹碚生.2002.黄瓜花性别分化和内源激素的关系.植物生理学通讯,38(4):317-320.
桂仁意,曹福亮,沈惠娟,谢寅峰.2003.多胺代谢对石竹试管苗成花中内源激素含量的影响.南京林业大学学报:自然科学版,27(4):27-30.
郝建军,刘延吉.2001.植物生理学实验技术.沈阳:辽宁科学技术出版社.
李秉真,孙庆林,张建华,马青枝.2000.苹果梨花芽分化期内源激素含量的变化(简报).植物生理学通讯,36(1):27-29.
李计红.2006.甜瓜性别分化的生理生化特性研究〔硕士论文〕.兰州:甘肃农业大学.
汪俏梅,曾广文.1997.苦瓜性别分化的激素调控.浙江农业大学学报,23(5):551-556.
王廷芹,杨暹.2008.外源赤霉素对青花菜茎尖内源激素含量的影响.中国蔬菜,(7):22-25.
肖华山,吕柳新,陈志彤.2006.荔枝花芽分化过程中多胺、核酸和蛋白质的动态.应用与环境生物学报,12(5):640-642.
徐继忠,陈江海,李晓东,张志华,王艳辉.2004.外源多胺对核桃雌雄花芽分化及叶片内源多胺含量的影响.园艺学报,31(4):437-440.
张多娇,齐红岩,陈璐璐.2009.嫁接对薄皮甜瓜花芽分化和花发育的影响.中国蔬菜,(6):25-30.
张新生,陈湖,于丽辰,傅友.2005.多胺与板栗花性别分化的关系.河北农业科学,4(9):5-9.
张建农,李计红.2007.甜瓜不同性别花蕾发育中激素和多胺的变化.园艺学报,34(5):1195-1200.
Altamura M M,Tomassi M.1998.Auxin,photoperiod and putrescine affect flower neoformation in normal androlB-transformed tobacco thin cell laye.Plant Physiology and Biochemistry,36(6):441-448.
de Dios P,Matilla A J,Gallardo M.2006.Flower fertilization and fruit development prompt changes in free polyamines and ethylene in damson plum(Prunus insiticaL.).J Plant Physiol,16:386-397.
Seo S G,Kang S W,Shim I S,Kim W,Fujihara S.2009.Effects of various chemical agents and early ethylene production on floral senescence ofHibiscus syriacusL.Plant Growth Regul,57:251-258.
