酵母双杂交筛选与转录因子ZNF580相互作用的蛋白*
2012-01-30罗玉玉孔麟麟任党丽张文成
罗玉玉, 杨 蓉, 孔麟麟, 任党丽, 张文成△
(1中国人民武装警察部队后勤学院生理学与病理生理学教研室,天津300162;2武警沈阳指挥学院门诊部,辽宁沈阳110034)
动脉粥样硬化(atherosclerosis,AS)是缺血性心脑血管疾病的重要病理基础[1],在我国发病率呈逐年上升趋势。AS是一种环境因素与遗传因素共同作用而诱发的多基因病,深入研究与AS相关基因的功能及其参与AS发生的机制是心脑血管疾病研究的热点领域。
ZNF580是由本课题组以致动脉粥样硬化因素——低密度脂蛋白(low-density lipoprotein,LDL)诱导人脐静脉内皮细胞系,筛选人主动脉cDNA文库,得到的一个新基因(Gen-Bank注册号:AF184939)。该基因编码172个氨基酸组成的蛋白质。经分子生物学网站进行蛋白质的功能分析显示:其N端为脯氨酸富含域,C端含有3个串联重复的C2H2型锌指蛋白结构域,可能为C2H2型锌指核转录因子家族的新成员[2-3]。最新的研究表明:C2H2型锌指核转录因子可能是真核细胞转录机制最重要的参与者,它们既可以结合DNA,又可以结合蛋白质,进而产生广泛的生物学效应[4-6]。为了寻找可能与ZNF580存在相互作用的蛋白,及其可能参与的细胞内信号转导通路,并最终揭示ZNF580的功能,本研究拟采用酵母双杂交技术钓取可能与ZNF580存在相互作用的蛋白。
材料和方法
1 菌株和质粒
Y190酵母菌购自Clontech;E.coli DH5α菌本室保存;酵母表达载体pGB-Vector购自上海睿星基因公司;人胎脑cDNA文库(载体为pACT2)购自上海睿星基因公司;pEGFPC1-ZNF580本室保存。
2 主要试剂和试剂盒
营养缺陷培养基购自Clontech;5-溴-4-氯-3-吲哚-β-D-半乳糖苷(5-bromo-4-chloro-3-indolyl β-D-galactoside,X-Gal)购自鼎国生物技术公司;鲑鱼精DNA购自Sigma;SfiⅠ限制性内切酶购自鼎国生物技术公司;T4 DNA连接酶购自鼎国生物技术公司;高保真Pfu DNA聚合酶购自Promega;质粒提取试剂盒(包括小量提取、大量制备2种)购自碧云天生物技术公司;DNA胶回收试剂盒购自Promega;PCR产物回收试剂盒购自Promega。
3 诱饵质粒构建
ZNF580扩增:以pEGFP-C1-ZNF580为模版,生工合成的ZNF580特异性引物对进行 PCR扩增(引物:5'-AAAAAGATCTTGCCCGGAGTGCGCCCGTG - 3',3'- AAAAAGCTTGTGGAGGCGCACGTGCTG-5')。PCR反应条件:94℃预变性3 min,94 ℃ 30 s,58 ℃ 30 s,72 ℃ 10 min,共30个循环。PCR产物过柱回收并酶切:反应按试剂盒说明书进行。目的片段回收并与载体连接:反应按试剂盒说明书进行。重组载体鉴定:重组载体转化细菌感受态DH5α,涂卡那霉素阳性平板37℃过夜。挑选7个单克隆摇菌过夜培养,提取质粒,用质粒作模板PCR扩增,产物行琼脂糖凝胶电泳,选择其中2个样品送TaKaRa测序。
4 诱饵蛋白自激活检测
采用LiAC转化法将诱饵质粒转化入酵母菌Y190。检测诱饵蛋白对LacZ报告基因是否存在转录激活作用。转化了诱饵质粒的酵母菌如果表达β-半乳糖苷酶,则可使底物X-Gal变蓝,说明存在自激活,否则说明无自激活。
5 酵母双杂交筛选人胎脑文库
5.1 人胎脑cDNA文库转化诱饵酵母菌 转库效率计算:转库效率只有达到1×107~1×108cfu/g DNA以上才能进行文库的筛选,以保证不会丢失阳性克隆。对生长在SD/-Trp/-Leu平板上的克隆计数,挑选克隆数在30~300之间的平板,根据稀释度计算转库效率。
5.2 营养缺陷平板初步筛选 人胎脑cDNA文库转化诱饵酵母菌,转化方法同上(LiAC转化法)。取200 μL菌液,涂布于150 mm SD/-Trp/-Leu/-His/+30 mmol/L 3-氨基 -1,2,4-三唑(3-amino-1,2,4-triazole,3-AT)平板上。第3 d起注意观察平板酵母克隆生长状态。菌在SD/-Trp/-Leu/-His+30 mmol/L 3-AT平板上生长7~14 d后,会有阳性克隆长出,阳性克隆一般大于3 mm。
5.3 β-半乳糖苷酶克隆转移滤纸实验进一步筛选 上述克隆全部进行β-半乳糖苷酶克隆转移滤纸实验,变蓝色的为阳性克隆。
5.4 一对一验证 随机挑取部分阳性克隆,抽提质粒,一对一转化诱饵酵母菌验证,阳性者抽提质粒送TaKaRa测序。
6 生物信息学分析
利用 NCBI网上数据库(http://www.ncb i.nlm.nih.gov)对返回的序列进行BLAST比对分析。
结 果
1 成功构建诱饵质粒
成功将ZNF580全长编码序列构建到酵母表达载体pGB上,见图1~4,说明pGB-ZNF580构建成功。
2 诱饵蛋白自激活检测
转化了诱饵质粒的酵母菌落未变蓝,转化了阳性对照质粒的酵母菌落变蓝,说明诱饵蛋白无自激活。可以进行下一步的筛库工作,见图5。

Figure 1.ZNF580 PCR product electrophoresis.图1 ZNF580 PCR产物电泳
Figure 2.ZNF580 PCR product electrophoresis after restriction enzyme digestion.图2 ZNF580 PCR产物酶切后电泳
Figure 3.The recombinant plasmid pGB-ZNF580 verified by PCR.M:marker;Lane1~7:PCR products.图3 重组质粒pGB-ZNF580 PCR鉴定

Figure 4.The recombinant plasmid pGB-ZNF580 verified by restriction enzyme digestion.Lane 1 and 2:digestion products.图4 重组质粒pGB-ZNF580酶切鉴定

Figure 5.Bait protein self-autoactivation assay.Left:the colonies of Y190 transformed with pCL1(positive control)turned blue after 2 h,which means the activation of lacZ reporter.Right:the colonies of Y190 transformed with pGB-ZNF580 didn't turn blue,which means the bait proteins had no self-autoactivation and could be used in library screening.图5 诱饵蛋白自激活检测

Figure 6.Thirty-five positive colonies identified by β -galactosidase colony-lift filter assay in yeast two-hybrid screening.图6 35个阳性克隆
3 人胎脑cDNA文库筛选
首先在营养缺陷平板上初步筛选,共获得169个阳性克隆,此时的1个酵母克隆可能含有2种或2种以上的文库质粒,把全部169个克隆分别在-Trp-Leu平板上划线分离单克隆后,再挑取单克隆在-Trp-Leu-His平板上培养,把培养板上的克隆全部进行β-半乳糖苷酶克隆转移滤纸实验进一步筛选获得136个阳性克隆。随机挑选其中40个阳性克隆从酵母中提取质粒,再把得到的质粒转入大肠杆菌DH5α中再次用氨苄西林抗性筛选,从中再次提取质粒,这样我们就获得了文库质粒,把获得的文库质粒一对一单独转入诱饵酵母细胞中,进行重复鉴定。只有转化了分离所得文库质粒后变蓝,转化了空载体后不变蓝的确认为最后的阳性克隆,最终获得35个阳性克隆,见图6,提取质粒,进行测序分析。
4 生物信息学分析
对35个阳性克隆相应的AD质粒进行测序,其中1个没有测出信号。对其余34个序列进行生物信息学分析,这些序列在GenBank数据库中是14种不同的基因。编码14种已知蛋白。其中有2种基因含有完整的编码序列,分别为SH3-domain GRB2-like 3(SH3GL3)和 D4,zinc and double PHD fingers family 1(DPF1);有12种基因含有部分编码区,分别为:activity-dependent neuroprotective protein(ADNP)、Smad family member 2(SMAD2)、glyoxalase domain containing 4(GLOD4)、kinesin family member 3A(KIF3A)、kinesin - associated protein 3(KIFAP3)、phospholipase A2- activating protein(PLAA)、phospholipase C gamma 1(PLCG1)、XPA binding protein 2(XAB2)、tripartite motif- containing 32(TRIM32)、suppressor of Ty16 homolog(SUPT16H)、Wilms tumor 1 interacting protein(WTIP)和 DIP2 disco-interacting protein 2 homolog C(DIP2C)。在这些序列中有部分重复,为同一个基因的相同或不同片段。
讨 论
本研究应用酵母双杂交技术筛选人胎脑cDNA文库,最终获得了14种可能与ZNF580存在相互作用的蛋白,并根据GenBank数据库及相关文献获得了它们的功能信息。这里我们重点讨论ZNF580与其中一种蛋白——磷脂酶C的相互作用关系。
磷脂酶C(phospholipase C,PLC)是普遍存在于哺乳动物细胞中的磷脂酰肌醇信号通路的关键酶。目前在哺乳动物中已鉴定出12种PLC同工酶,根据其不同的氨基酸序列分为5型:PLC-β、PLC-γ、PLC-δ、PLC-ε和 PLC-ξ。许多细胞外信号分子,如激素、神经递质、免疫球蛋白、细胞因子、生长因子等都可以激活磷脂酰肌醇特异性PLC。进而将信息传递给下游效应分子,调节细胞的生命活动[7]。研究发现,PLC-γ1不仅具有催化分解磷脂酰肌醇二磷酸(PIP2)的脂酶功能,而且也具有蛋白接合器的作用,PLC-γ1分子除了由X和Y区构成的酶活性部位外,还有作为蛋白质相互结合的SH(Src homology domain)区,为2个SH2结构域和1个SH3结构域。通过这些结构区域与其它信号转导分子结合,产生多种细胞反应[8-9]。目前已知的可以和 PLC-γ1相互作用的蛋白有 Rac1、PLD2、Grb2、Akt等[10-12]。
PLC-γ1是许多能自身磷酸化的生长因子受体作用的底物。如血小板生长因子、表皮生长因子、成纤维细胞生长因子、胰岛素生长因子等[13]。PLC-γ1的活化主要通过蛋白酪氨酸激酶(protein tyrosine kinase,PTK)途径。多种生长因子与具有内源性PTK活性的受体型PTK结合后,受体发生二聚化,导致受体内在的PTK激活,使受体自身的酪氨酸残基磷酸化,产生对PLC-γ1具有高亲和力的结合位点,活化的PTK能特异性地识别PLC-γ1的SH2结构域,并将蛋白质序列中的酪氨酸磷酸化,从而激活PLC-γ1。激活的PLC-γ1水解PIP2产生二酰甘油(DAG)和三磷酸肌醇(IP3),DAG能激活蛋白激酶C,IP3能引起细胞内钙库的释放,通过这两条通路或者和其它未知通路形成广泛的crosstalk,从而启动多种细胞反应,影响着细胞的生长、分化、增殖、凋亡等功能[14]。
本课题组前期对ZNF580蛋白分子进行生物信息学分析表明:ZNF580蛋白分子上存在一系列的蛋白酶磷酸化作用位点,其中141~143位THR氨基酸为蛋白激酶C磷酸化作用位点;130~133位KRSS氨基酸为cAMP及cGMP依赖型蛋白激酶磷酸化作用位点。可以推测,ZNF580在细胞内作为核转录因子的活性受磷酸化作用的调节。如果ZNF580与PLC-γ1之间存在相互作用,那么ZNF580很可能参与PTK途径的信号转导,其可能的信号通路见图7。
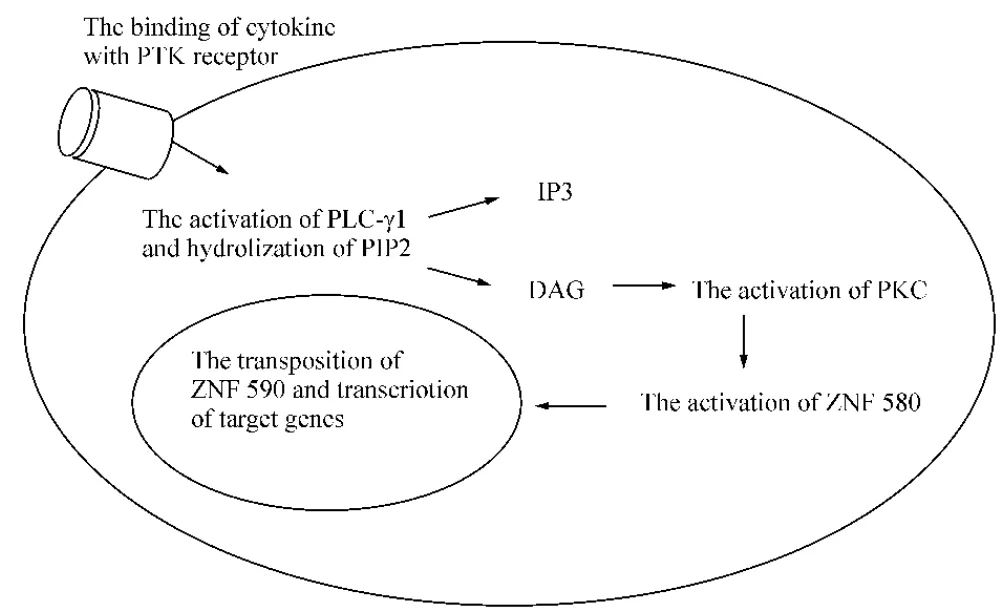
Figure 7.The possible mechanism of ZNF580 involved in protein tyrosine kinase(PTK)signal pathway.PLC:phospholipase C;PIP2:phosphatidylinositol biphosphate;IP3:inositol triphosphate;DAG:diacylglycerol;PKC:protein kinase C.图7 ZNF580参与PTK信号通路的可能机制
但是关于ZNF580参与的信号转导途径及其功能的研究目前仍然处于起步阶段,而且由于蛋白质分子间的相互作用非常复杂,时空特异性等方面的偏差会导致一些人为的假象。酵母细胞是最简单的真核细胞,其体内蛋白质合成后加工修饰类型及其程度与哺乳动物的相应过程存在一定差别,因此酵母双杂交的结果仅仅是初步提出相互作用的可能性,还需要进一步的实验证实。
[1]Lusis AJ. Atherosclerosis [J]. Nature,2000, 407(6801):233-241.
[2]张文成,陈保生,曾武威,等.低密度脂蛋白诱导下调的新基因cDNA的克隆及组织表达[J].基础医学与临床,2003,23(3):279-282.
[3]张文成,孙慧燕,孟祥艳,等.ZNF580-EGFP融合蛋白的亚细胞定位研究[J].中国病理生理杂志,2008,24(8):1590-1594.
[4]Wang J,Kudoh J,Takayanagi A,et al.Novel human BTB/POZ domain-containing zinc finger protein ZNF295 is directly associated with ZFP161[J].Biochem Bioph Res Commun,2005,327(2):615-627.
[5]Al-Kandari W,Koneni R,Navalgund V,et al.The zinc finger proteins ZXDA and ZXDC form a complex that binds CIITA and regulates MHC II gene transcription[J].J Mol Biol,2007,369(5):1175 -1187.
[6]Mysliwiec MR,Kim TG,Lee Y.Characterization of zinc finger protein 496 that interacts with Jumonji/Jarid2[J].FEBS Lett,2007,581(14):2633 -2640.
[7]Rliee SG,Choi KD.Regulation of inositol phospholipids specific phospholipase C isozymes[J].J Biol Chem,1992,267(18):12393-12396.
[8]Carpenter G,Ji QS.Phospholipase C - γ as a signal transducing element[J].Exp Cell Res,1999,253(1):15-24.
[9]Rebecchi MJ,Pentyala SN.Struction,function,and control of phosphoinositide-specific phospholipase C[J].Physiol Rev,2000,80(4):1291-1335.
[10]Li S,Wang Q,Wang Y,et al.PLC - γ1 and Rac1 coregulate EGF-induced cytoskeleton remodeling and cell migration[J].Mol Endocrinol,2009,23(6):901 -913.
[11]Jang IH,Lee S,Park JB,et al.The direct interaction of phospholipase C - γ1 with phospholipase D2 is important for epidermal growth factor signaling[J].J Biol Chem,2003,278(20):18184-18190.
[12]Cho JH,Hong WP,Yun S,et al.Grb2 negatively regulates epidermal growth factor-induced phospholipase C - γ1 activity through the direct interaction with tyrosine-phosphorylated phospholipase C - γ1[J].Cell Signal,2005,17(10):1289-1299.
[13]Ye K,Aghdasi B,Luo HR,et a1.Phospholipase Cγ1 is a physiological guanine nucleotide exchange factor for the nuclear GTPase PIKE[J].Nature,2002,415(6871):541-544.
[14]Anderson D,Koch CA,Grey L,et a1.Binding of SH2 domains of phospholipase C gamma 1,GAP,and Src to activated growth factor receptors[J].Science,1990,250(4983):979-982.
