肝细胞生长因子对喉返神经损伤后再生的作用*
2012-01-12李孟陈世彩陈东辉夏思文李晓雨郑宏良
李孟 陈世彩 陈东辉 夏思文 李晓雨 郑宏良
喉返神经损伤后,再生的轴突数目不足及功能恢复不佳是影响喉功能恢复的重要原因。应用神经营养因子等生物活性因子促进神经轴突的再生,近年来已成为周围神经损伤修复研究中的一项重要突破。肝细胞生长因子(hepatocyte growth factor,HGF)广泛分布于全身各组织器官,是一种多效性细胞因子,具有多种功能。已有研究证实HGF对中枢神经系统有保护作用[1],在坐骨神经[2]、舌下神经[3]及面神经[4]等周围神经损伤后,局部给予外源性HGF对神经元有保护作用并对轴突再生和诱向生长具有促进作用。HGF对喉返神经损伤后修复的作用尚未见相关报道。本研究通过制造犬单侧喉返神经损伤动物模型后,用聚乳酸聚羟基乙酸共聚物[Poly(DL lactide co glycolide),PLGA]管桥接喉返神经断端,并在局部应用外源性HGF,观察其对受损喉返神经再生及功能恢复的作用,报告如下。
1 材料与方法
1.1实验动物与分组 健康成年杂种犬20只,体重8.0~15.0 kg,雌雄不拘,购于第二军医大学实验动物中心。首先制作单侧喉返神经损伤动物模型:于第4气管环水平、相当于环状软骨下缘下2 cm处切断实验犬左侧喉返神经并切除一段长约5 mm的神经,用11-0无创缝线套接神经断端入8 mm长的PLGA管中,神经断端间距约5 mm;颈部皮瓣缝合前,测量桥接管与体表的距离;然后,将实验犬分为HGF组和对照组,每组10只。HGF组术中用微量注射器将HGF (50 ng/μl)10 μl注入PLGA管内,对照组同法注射等量的生理盐水。HGF组和对照组术后隔日分别在相当于神经桥接处经皮局部注射HGF 0.1 ml或等量生理盐水1次,共1个月,注射深度以术中测量的结果为依据。所有动物的右侧喉返神经不予处理,作为正常对照。
1.2电子喉镜观察声带运动 分别于两组动物术前、术后即刻、术后2、3、4个月时行电子喉镜检查,在平静呼吸及麻醉变浅时,捏鼻刺激诱犬发声,观察双侧声带的形态、运动及声门闭合情况。
1.3喉返神经及喉肌的电生理检查 每次喉镜检查后,按先前报告的方法[5],以四导同心针电极记录动物双侧环杓后肌及甲杓肌在发音及呼吸状态下的自发肌电位,并以波宽0.1 ms、电流强度为40~80 mA的电流在犬胸骨上窝、气管食管沟间经皮肤刺激左侧喉返神经桥接处近心端神经及对侧喉返神经相同平面,记录双侧甲杓肌的自发肌电位幅度、最大诱发电位幅度及其潜伏期,术侧的自发肌电位幅度、最大诱发电位幅度及其潜伏期以其占该动物健侧的比例即恢复率(%)表示。
1.4再生的喉返神经光镜及超微结构观察 完成最后一次神经电生理检查后,即刻处死动物。取神经桥接处远端神经干1 cm, 常规戊二醛固定,1%锇酸后固定,梯度脱水,Epon812包埋,半薄切片,1%甲苯胺蓝染色,在显微镜下用图像分析系统计算再生有髓神经纤维轴突数目、直径及髓鞘厚度[6]。超薄切片,H-800透射电镜下观察再生神经干的形态。
1.5统计学方法 用SPSS13.0软件对两组上述结果进行两个独立样本t检验。
2 结果
实验过程中因为疾病、麻醉等原因5只犬死亡,其中HGF组死亡2只,对照组死亡3只。
2.1电子喉镜观察 术后2个月时两组动物术侧声带均明显缩短、固定于旁正中位,发声时声门有明显裂隙。术后3个月时HGF组有4只犬声带恢复内收及外展运动,但幅度较小;而对照组动物仍未恢复运动。术后4个月时,HGF组存活的8只犬中有7只术侧声带恢复内收、外展运动,运动幅度均较大,1只犬术侧声带仍固定于旁正中位;对照组存活的7只犬中5只犬术侧声带恢复内收及外展运动,但幅度小于HGF组,另2只犬术侧声带固定于旁正中位。除3只未恢复运动的犬发声时声门后联合有裂隙外,其余犬声门裂隙不明显,两组犬健侧声带活动均正常。
2.2神经电生理学检查结果 术后2个月时两组犬即可记录到喉肌的自发与诱发电位,且HGF组的自发及诱发电位均明显大于对照组,差异均有统计学意义(P<0.05)。术后3个月时,二组犬的喉肌自发及诱发电位均明显增强,潜伏期明显缩短,与同组术后2个月时比较,差异均有统计学意义(P<0.05);HGF组三项观察指标均明显优于对照组,差异均有统计学意义(P<0.05)。术后4个月时两组的自发及诱发肌电位均进一步增大,但与同组3个月时比较,差异无统计学意义(P>0.05),潜伏期进一步缩短,与同组的3个月比较差异均有统计学意义(P<0.05)。尽管如此,术后4个月时HGF组的三项神经电生理学观察指标均明显优于对照组,差异均有统计学意义(P<0.05)(表1)。
2.3再生神经纤维形态学观察 HGF组神经桥接处远端的神经干内再生神经纤维数量明显多于对照组,有髓神经纤维的直径、髓鞘厚度均明显大于对照组,差异均有统计学意义(P< 0.05),但再生的神经纤维数量仍然明显少于健侧喉返神经,有髓神经纤维的直径、髓鞘厚度均明显小于正常喉返神经,差异有统计学意义(P<0.05)。透射电镜下显示HGF组再生神经纤维直径、髓鞘厚度以及同一放大倍数视野下神经纤维数量均明显大于对照组,形态结构接近正常,而对照组形态结构与健侧神经比较差异较大,可见较多的无髓神经纤维(表2,图1)。
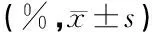
表1 两组动物术后不同时间神经肌电图检查结果
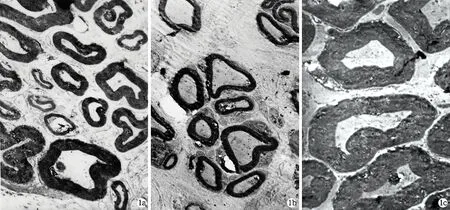
图1 神经干电镜图
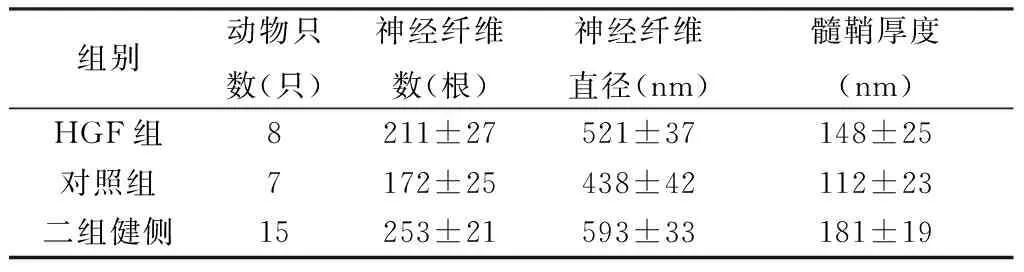
组别动物只数(只)神经纤维数(根)神经纤维直径(nm)髓鞘厚度(nm)HGF组8211±27521±37148±25对照组7172±25438±42112±23二组健侧15253±21593±33181±19
3 讨论
喉返神经损伤致声带麻痹是临床上常见的疾病之一,尽管喉返神经修复治疗单侧声带麻痹已取得令人鼓舞的效果[7],但长期损伤修复后的神经功能恢复仍不理想。神经轴突的生长能力主要依赖于轴突周围的微环境而非其内在特性,而神经营养因子、神经趋化生长因子等是构成周围微环境的主要来源[8]。研究证实,应用外源性生物因子,如睫状神经生长因子(CTNF)、神经生长因子(NGF)、白细胞介素-1(IL-1)等,可促进神经轴突的生长,并有助于维持运动神经元的功能[9~11]。但上述生物活性因子对神经轴突和神经元保护作用的局限性导致其单独应用不能取得理想的效果,因为长期失神经后的肌肉萎缩和纤维化使得神经、肌肉再生的微环境不理想,从而阻碍了神经功能的理想恢复。
HGF最早于1984年从肝切除术后的残余肝组织中发现,因其可刺激肝细胞合成DNA而命名为肝细胞生长因子。近来发现,HGF还是一种多效性神经营养因子,在全身多处均有表达[12],在神经系统的生长发育过程中有促进作用[13],并对运动神经元的发育、存活、轴突的生长和导向、肌肉的营养均有重要作用[14~16]。在外周神经系统损伤或被切断后,内源性HGF及其受体c-Met 表达均增加,对神经元有保护作用,对轴突有促生长作用,并能促进神经纤维的修复[17]。Hashimoto等[2]发现大鼠坐骨神经切断模型中有HGF及其受体的表达,认为HGF/c-Met参与调节坐骨神经损伤后的再生修复过程;冯凯等[18]发现HGF能促进大鼠坐骨神经损伤后的有髓纤维再生,并促进感觉、运动神经纤维恢复。还有研究证实外源性应用HGF对面神经损伤后神经干、神经元及肌运动终板都有保护作用,同时HGF对面肌还有营养作用[4,19]。唐金树等[20]研究认为HGF可以促进运动神经末梢在肌肉缝合界面的穿行,在失神经支配后的肌肉中形成更多的运动终板结构,因此应用外源性HGF能促进肌肉神经再生。
有研究表明,周围神经轴突切断后,运动神经元上的HGFmRNA表达水平并不升高,因此其产生的内源性HGF不足以维持运动神经元的功能[20]。提示在外周神经损伤时,应用外源性HGF甚为重要[21]。本研究通过切断犬一侧喉返神经,用PLGA管将两断端桥接,HGF组局部应用外源性HGF,术后2个月,尽管两组的声带运动恢复情况不明显,但电生理学检查结果显示HGF组均明显优于生理盐水对照组。而术后4个月,HGF组的声带运动恢复情况明显优于生理盐水对照组,且喉返神经肌电图自发电位与诱发电位幅度、潜伏期、再生神经轴突计数、神经纤维直径及髓鞘厚度方面均优于对照组,差异有统计学意义。由此证明,喉返神经切断后局部给予HGF对喉返神经再生修复具有促进作用。另外,骨骼肌的营养与再生是喉返神经损伤后神经修复治疗效果的重要决定因素之一。失神经支配使骨骼肌发生萎缩和纤维化,必需要有足够的再生种子细胞和良好的再生微环境才能保证肌肉再生并获得满意的功能恢复。研究表明,损伤骨骼肌细胞的表面均有HGF及其受体c-Met的表达[2],说明应用外源性HGF不仅能促进肌细胞的增殖及分化,同时能改善肌肉再生微环境(促进血供、抗凋亡、肌萎缩、肌纤维化),从而发挥协同作用,以促进失神经骨骼肌的再生,达到喉肌功能的良好恢复的效果。
外源性营养因子的给药途径和方法研究甚多。有效的神经营养因子浓度及持续作用时间可以更有效的维持运动神经元的存活,促进运动神经元轴突的再生[22]。为了在再生神经轴突周围的微环境中维持有效的神经营养因子浓度,有研究者将营养因子直接注入桥接神经的硅胶管内[23],但注入的神经营养因子有效的作用时间较短。也有研究者将营养因子注射至皮下、肌肉、腹腔等, 通过局部吸收,经过全身血液循环作用于局部的神经元、再生的神经轴突[24],但不足之处是易引起全身的药物不良反应。本研究在上述研究的基础上,通过桥接管内注入HGF, 同时隔日在桥接管周围注射HGF, 通过药物的弥散作用,达到营养因子直接作用于再生神经轴突,并通过轴突的吸收,作用于运动神经元。此种方法HGF也可能被局部组织吸收,通过全身血循环作用到再生的神经元、轴突及肌肉,从而进一步促进神经肌肉的再生,但全身吸收的副作用难以避免。因此,如何在局部通过HGF缓释作用于神经损伤部位,既保持局部稳定的药物浓度,又避免因药物全身吸收所带来的不良反应,有待于进一步研究。
4 参考文献
1 Tsuzuki N, Miyazawa T, Matsumoto K,et al. Hepatocyte growth factor reduces infarct volume after transient focal cerebral ischemia in rats[J]. Acta Neurochir Suppl, 2000,76: 311.
2 Hashimoto N, Yamanaka H, Fukuoka T, et al. Expression of HGF and cMet in the peripheral nervous system of adult rats following sciatic nerve injury[J]. Neuroreport, 2001,12:1 403.
3 Okura Y, Arimoto H, Tanuma N, et al. Analysis of neurotrophic effects of hepatocyte growth factor in the adult hypoglossal nerve axotomy model[J]. Eur J Neurosci, 1999,11: 4 139.
4 周翠英,骆文龙. 肝细胞生长因子对面神经损伤修复作用的实验初探[J].重庆医学,2003,32:299.
5 Zheng H, Zhou S, Chen S,et al. An experimental comparison of different kinds of laryngeal muscle reinnervation[J]. Otolaryngol Head Neck Surg, 1998,119: 540.
6 郑宏良,周水淼,李兆基,等. 睫状神经营养因子在喉返神经再生过程中的表达及分布[J].中华耳鼻咽喉科杂志,1999,34:289.
7 Zheng H, Li Z, Zhou S,et al. Update: laryngeal reinnervation for unilateral vocal cord paralysis with the ansa cervicalis[J]. Laryngoscope, 1996,106: 1 522.
8 易声禹,张剑宁. 神经组织移植与神经再生的历史回顾[M].北京: 人民卫生出版社, 1993.43~62.
9 郑宏良,何成,周水淼,等. 外源性睫状神经生长因子对喉神经再生的趋向作用[J]. 中华耳鼻咽喉科杂志, 2002,37:333.
10 黄立,洪光祥,王发斌.神经生长因子对周围神经端侧吻合靶器官功能恢复影响的实验研究[J]. 中国临床康复,2004,8:1 254.
11 宫可同,阚世廉,鲁毅军. 白细胞介素-1在周围神经损伤后神经再生中的作用[J]. 中华手外科杂志,2003,19:124.
12 Wolf HK, Zarnegar R, Michalopoulos GK. Localization of hepatocyte growth factor in human and rat tissues: an immunohistochemical study[J]. Hepatology, 1991. 14: 488.
13 Gutierrez H, Dolcet X, Tolcos M, et al. HGF regulates the development of cortical pyramidal dendrites[J]. Development, 2004. 131: 3 717.
14 Koyama J, Yokouchi K, Fukushima N, et al. Neurotrophic effect of hepatocyte growth factor on neonatal facial motor neurons[J]. Neurol Res, 2003. 25: 701.
15 Caton A, Hacker A, Naeem A, et al. The branchial arches and HGF are growth-promoting and chemoattractant for cranial motor axons[J]. Development, 2000. 127: 1 751.
16 Krull CE, Koblar SA. Motor axon pathfinding in the peripheral nervous system[J]. Brain Res Bull, 2000,53: 479.
17 周翠英,骆文龙. 肝细胞生长因子对周围神经再生的作用研究[J].药学专论,2002,11:42.
18 冯凯,林世和,吕宪民,等. 肝细胞生长因子对大鼠坐骨神经损伤保护作用的实验研究[J]. 中国神经免疫学和神经病学杂志,2005,12:122.
19 刘阳,苏俊波,骆文龙. 肝细胞生长因子对面神经损伤后运动终板及肌纤维修复作用的研究[J]. 重庆医学,2004,33:1 231.
20 唐金树,刘郑生,赵斌,等. 肝细胞生长因子对肌肉水平神经再生影响的初步研究[J].中国临床康复,2004,8:4 448.
21 Boros P, Miller CM. Hepatocyte growth factor: a multifunctional cytokine[J]. Lancet, 1995,345: 293.
22 于雯雯,蒋欣泉,张志愿. 周围神经导管修复中神经营养因子的应用方式进展[J]. 中国口腔颌面外科杂志,2009, 1:77.
23 Gillespie LN. Regulation of axonal growth and guidance by the neurotrophin family of neurotrophic factors[J]. Clin Exp Pharmacol Physiol, 2003,30: 724.
24 刘鸿飞, 徐勇忠. CNTF 对端侧吻合后面神经再生作用的实验研究[J].口腔医学研究,2002,18: 251.
