中国刺参(♂)×日本红刺参(♀)杂交子代的胚胎、幼体发育及幼参生长性状的研究*
2012-01-05孙秀俊孔令锋于瑞海
孙秀俊,李 琪,孔令锋,于瑞海
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
中国刺参(♂)×日本红刺参(♀)杂交子代的胚胎、幼体发育及幼参生长性状的研究*
孙秀俊,李 琪**,孔令锋,于瑞海
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
本研究建立中国刺参(♂)与日本红刺参(♀)杂交组合,通过对杂交子代的胚胎发育进行观察,详细描述其胚胎及幼体各个阶段的发育时序和形态特征,并比较在幼参培育期间杂交苗与中国刺参自交苗的生长和成活。结果表明,在20~22℃时,杂交子代发育至初耳幼体的时间为受精后24 h 38 min,发育至中耳幼体的时间为72 h 8 min,然后经过167 h 8 min(约7d)发育至大耳幼体,到受精后第9天发育至樽形幼体。樽形幼体后期,纤毛环逐渐退化,浮游能力减弱,幼虫由浮游转至底栖生活。在幼参培育期间,受精后第38~51天,平均水温为25.2℃,杂交幼参的体重特定生长率(SGR)为2.58%,而自交幼参的SGR略高,为3.03%;从受精后第51~80天,平均水温达到27.17℃,杂交幼参的生长速度减慢,SGR减小至0.66%,但平均体质量明显增加,到第80天达到(0.34±0.035)g,而自交幼参活动能力减弱,摄食量明显减少,SGR减小至0.08%,生长近于停滞,到第80天平均体质量为(0.19±0.034)g。自交苗和杂交苗的成活率相差不显著,分别为82.2%和73.9%。这表明,在高温条件下杂交苗种比自交苗种生长优势明显。
刺参;杂交;胚胎发育;生长
刺参(Apostichopus japonicus)隶属于棘皮动物门(Echinodermata)、海参纲(Holothuroidea)、楯手目(Aspidochirota)、刺参科(Stichopodidae),主要分布在我国的辽宁、山东、河北等沿海省,朝鲜半岛、日本和俄罗斯远东沿海,是我国重要的特种海洋经济动物。近年来,随着海参市场需求的不断增长,我国刺参养殖呈现迅猛发展态势,养殖产量不断增加,据统计,2008年我国刺参总产量已达到92 567 t,其中超过95%来自山东和辽宁2个省[1]。然而,随着生产规模的扩大、产量的增加,却出现了苗种质量下降、养殖刺参生活力降低和疾病频发等问题,尤其是在高温季节幼参生长停滞、死亡惨重,严重制约了海参养殖业的健康发展。因此,进行刺参的遗传改良、培育高产抗逆新品种,对我国刺参养殖产业和区域经济的可持续发展愈显重要。
杂交育种仍是目前国内外动植物遗传育种中应用最广泛、成效最显著的育种方法之一,也是水产动物育种的重要途径之一。由于杂种优势的存在,杂交能迅速和显著地提高杂种的产量或生活力,因此,利用杂种优势获得水产动物的生长、存活等表型性状改良的研究,国内外均有报道。目前杂交育种已广泛应用于鱼类[2-5]和贝类[6-10]等水产动物中的育种实践,并且在提高生长速度、抗病力、抗逆性、出肉率、成活率、饵料转化率和改良肉质等方面取得了一定的成效。杂交工作包括种间杂交和种内杂交,对于海洋无脊椎动物,种间杂交往往会由于亲缘关系相距较远,而出现杂交不亲和、幼体不成活和杂种后代育性差等难以解决的问题,而种内杂交不存在此类问题,可操作性强,容易推广,是杂种优势利用的主要途径[11]。
在日本,野生刺参存在红刺参、黑刺参和青刺参3个品系。Kanno等[12-13]分别采用同工酶标记和微卫星分析了3种体色刺参的遗传差异,结果显示红刺参与其他两色刺参之间存在明显的遗传分化,并且推测可能存在生殖隔离。Sun等[14]通过比较3种颜色刺参的线粒体全基因组,证明三者在分类地位上属于同1个种。红刺参肉体较厚,市场价格较高,一般为青、黑刺参的2~3倍。鉴于其优良的经济价值,2006年中国海洋大学从日本成功引进红刺参新品种,开展了红刺参和中国刺参的近缘杂交技术研究,胡美燕和李琪[15]报道了日本红刺参♀×中国刺参♂杂交组在幼虫期和稚参期的各生长性状平均值均大于其它3组杂交和自交组合,杂种优势率均较为明显。本研究采用此最优杂交组合,以红刺参为母本,中国刺参为父本,对杂交子代胚胎发育各个阶段的发育时序和形态特征进行详细的描述,并比较在高温条件下杂交幼参和中国刺参自交幼参的生长和成活,以期为刺参杂交苗种培育提供基础资料。
1 材料与方法
日本红刺参采自日本大分县,中国刺参采自我国乳山沿海。以中国刺参为父本,红刺参为母本进行杂交。亲参蓄养和苗种的培育均在海阳市海泰育苗场进行。采用阴干升温法诱导种参产卵,亲参阴干1 h,流水刺激40 min,随后将种参放入升温海水中(较原亲参蓄养水温升高3~5℃),待其排精、产卵。2009年4月27日收集红刺参的卵子和中国刺参的精子,混合受精,建立杂交实验组;同时收集中国刺参的精子和卵子,混合受精后建立中国刺参自交实验组。幼体培育在50 L的塑料水桶中进行,培育密度为0.5个/m L,培育初期的水温维持在20~22℃,海水盐度30~32,当自然海水温度达到20℃以上时,采用常温海水进行培育。
浮游幼体培育期间,投喂饵料种类包括牟式角毛藻和小新月菱形藻,日投饵2次,投饵量根据胃部饱满度和生长发育的情况而定。日换水2次,每次换水量为1/3~1/2,持续充气。定期随机抽样观察30个耳状幼体,目微尺测定体长。当20%~30%的耳状幼体发育至樽形时投放附着基。幼参培育期间投喂鼠尾藻、马尾藻和海带等人工配合饲料,日投饵2次,投饵量根据稚参摄食情况而定。日换水2次,每次换水1/3~1/2,根据池底粪便量和残饵量每隔7~10 d倒池1次。在受精后第38、51和80天测量幼参体质量,体重特定生长率(SGR)按照下面的公式计算:SGR(%·d-1)=100(ln W2-ln W1)/T。其中,W1和W2分别为幼参开始的体质量,T为持续的天数。
2 结果
2.1 自交苗和杂交苗胚胎和幼体发育过程中的外部形态特征
杂交子代和自交子代的胚胎及幼体发育时间见表1。图1和2分别为杂交子代和自交子代的发育图谱。中国刺参自交子代与杂交子代的胚胎和幼体发育时序完全相同,但是发育至各个阶段的时间略有不同。杂交子代发育至大耳幼体的时间为167 h 8 min,而自交子代发育至大耳幼体的时间略长,为211 h 32 min。杂交子代的变态附着比自交子代提前1 d左右的时间。现以杂交子代的胚胎及幼体发育为例,详细描述各个阶段的形态特征。
受精卵:受精卵为沉性卵,卵黄均匀透明,受精卵的平均卵径为(175.91±3.56)μm。受精后约30 min,出现第一极体(见图1-1)。
卵裂:卵子受精后52 min进入第1次细胞分裂。第1次分裂为纵裂,形成2个大小相等分裂球(见图1-2)。第2次也为纵裂,与第1次分裂面相垂直,产生4个大小相等的分裂球(见图1-3)。第3次为横裂,产生8个大小相等的分裂球(见图1-4)。受精卵经多次分裂,细胞数量不断增加,细胞体积越来越小(见图1-5)。
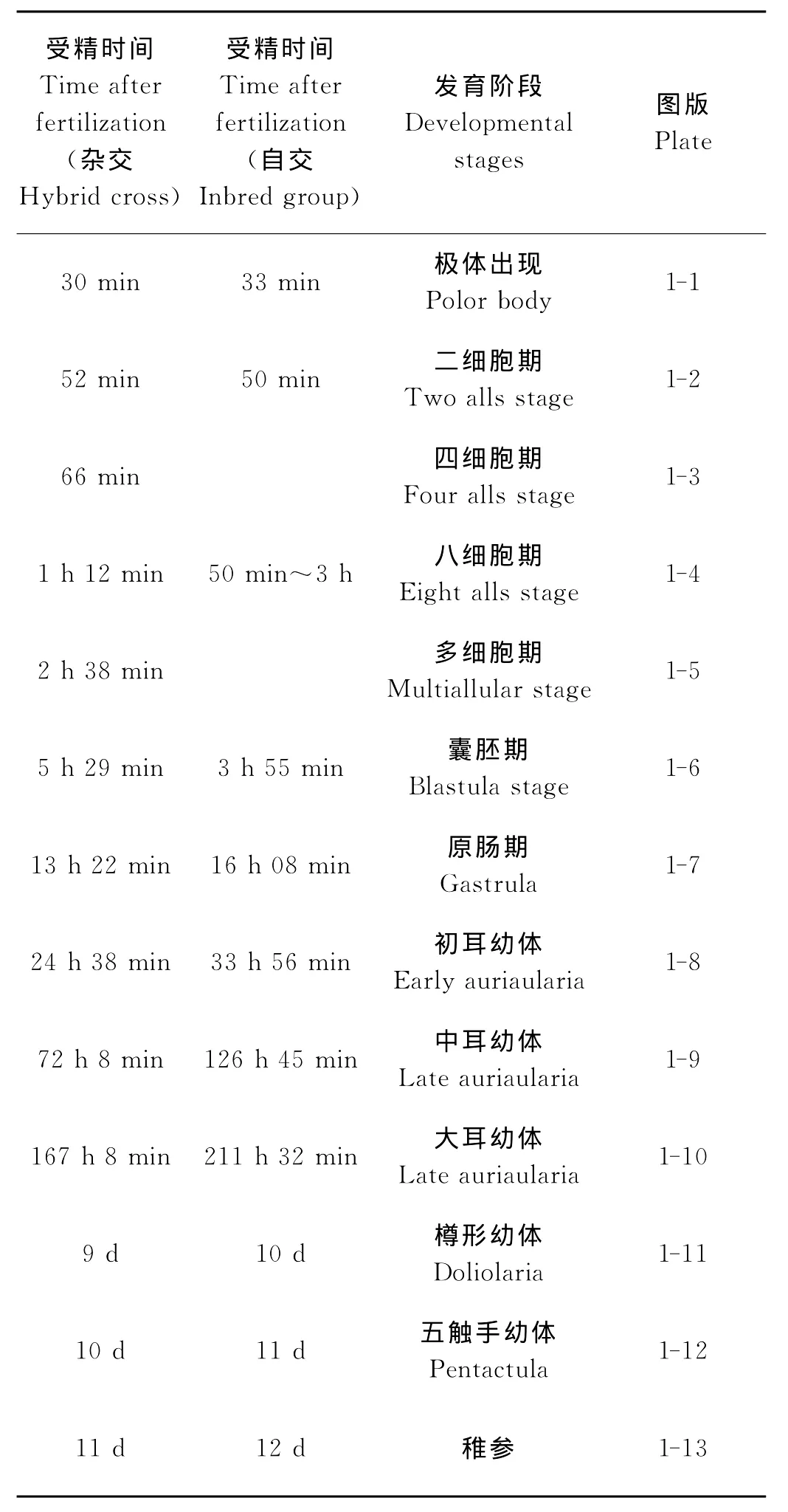
表1 中国刺参(♂)与日本红刺参(♀)杂交子代和中国刺参自交子代的胚胎及幼体发育(水温20~22℃)Table 1 Embryonic and larval development of the inbred and hybrid between Green(♂)and Red(♀)sea cucumber(Temperature:20~22℃)
囊胚期:受精后5 h 29 min,多次卵裂之后,细胞界限开始模糊,胚体中央出现1个大而且圆的空腔,形成囊胚腔(见图1-6)。
原肠期:受精后13 h 22 min,囊胚拉长,经过内陷后形成原肠腔,到原肠后期,原肠腔向胚体一侧倾斜,成为幼体腹侧,原口形成肛门(见图1-7)。
初耳幼体:受精后24 h 38 min,幼体臂形成,消化系统开始完善,可以明显区分口、食道、胃、肠和肛门等消化器官,原体腔和水体腔均未发育(见图1-8)。
中耳幼体:受精后72 h 8 min,幼体臂粗壮而且明显,体长明显增长,左侧原体腔开始延长,水体腔未延长(见图1-9)。
大耳幼体:受精后167 h 8 min,出现5对球状体,体长进一步增长,左侧原体腔进一步发育,长度超过胃长的一半,水体腔延长,到大耳后期,在水体腔外侧形成5个囊状五触手原基(见图1-10)。
樽形幼体:受精后9 d,耳状幼体不断收缩,纤毛带失去连续性,逐渐变为圆筒形,体长缩小为原来的一半左右,内部消化器官很难分辨,即发育至樽形幼体。到樽形后期,幼体纤毛运动能力减弱,活动缓慢,主要集中于水的中下层(见图1-11)。
五触手幼体:受精后10 d,5个触手伸出前庭,形成五触手幼体(见图1-12)。
稚参:受精后11 d,身体拉长,在幼体腹面左侧后端形成第一管足,随后管足数量不断增加,并在幼体体表形成许多钙质骨片,形态与成体海参基本相似(见图1-13)。

图1 中国刺参(♂)×日本红刺参(♀)杂交苗种的胚胎幼体发育和幼参生长Fig.1 Embryonic,larval development and juvenile growth of the hybrid between Green(♂)and Red(♀)sea cucumber

图2 中国刺参自交子代的胚胎幼体发育和幼参生长Fig.2 Embryonic,larval development and juvenile growth of the inbred Green

图3 自交子代和杂交子代幼体的体长生长Fig.3 Laval growth of the inbred and hybrid sea cucumber
2.2 浮游幼体的生长
随着培育天数增加,自交子代和杂交子代浮游幼体的体长不断增加,到受精后第10天均达到最大值,分别为(680.57±54.92)μm和(638.18±55.62)μm(见图3)。自交子代和杂交子代的体长生长率,在7 d分别为73.35和59.38μm/d;在10 d,由于幼体发育至樽形阶段,身体收缩,体长减小,分别为59.40和46.39μm/d。自交子代和杂交子代幼体的平均体长在7 d和10 d均差异不显著。
2.3 杂交幼参和自交幼参的生长及成活比较
由图4可知,受精后第38天,自交幼参和杂交幼参的平均体质量之间存在显著差异(P<0.05),杂交幼参的平均体质量为(0.20±0.07)g,自交幼参的平均体质量为(0.13±0.04)g。从第38~51天,水温保持在25℃左右(平均水温为25.2℃),杂交苗的体质量特定生长率(SGR)为2.58%,而当地自交苗的SGR略高,为3.03%,到第51天,两者之间的平均体质量差异不显著(P>0.05)。从受精后第51~80天,水温上升至27℃以上(平均水温为27.2℃),杂交苗的生长速度略有减慢,SGR减小为0.66%,但平均体质量明显增加,到第80天为(0.34±0.035)g,而在此期间,自交苗活动能力明显减弱,摄食量减少,SGR减小至0.08%,生长近于停滞,第80天平均体质量为(0.19±0.034)g,此时杂交幼参和自交幼参之间的平均体质量差异极显著(P<0.01)。
另外,随着培育天数的增加,自交苗和杂交苗的个体间体质量差异逐渐增大,尤其是杂交苗的体质量差异极为悬殊,在第80天时,最重的杂交苗个体为3.58 g,最轻的个体不足0.01 g;而最重的自交苗个体仅为0.77 g,最轻的个体也不足0.01 g。从受精后第38天到第80天,自交苗的成活率为82.2%,杂交苗成活率略低,为73.9%。第80天杂交幼参的背面观和腹面观见图1-14和图1-15。杂交幼参的背部呈黄褐色,偏向于中国刺参,腹部纵带为红色,偏向于红刺参,并且刺型优良,具有中国刺参和红刺参的共同特征。

图4 中国刺参(♂)×日本红刺参(♀)杂交幼参和中国刺参自交幼参的平均体质量Fig.4 Comparison of body weight of the hybrid(Green♂×Red♀)and inbred juveniles(Green)
3 讨论
水产动物育种中,利用种内不同地理种群杂交进行生长性状的改良,已应用于扇贝、鲍鱼和马氏珠母贝等水产养殖的重要经济种类,并取得了一定的进展[16-18]。相对于种间杂交,种内杂交不容易产生种质污染,有利于经济种类的种质资源保护。因此,为解决我国养殖刺参生活力降低、疾病频发等问题,种内杂交育种是培育刺参优良品种的重要途径之一。
本实验描述了中国刺参(♂)与日本红刺参(♀)的种内杂交后代的胚胎及幼体各个阶段的发育时序和形态特征。杂交子代受精卵的卵裂期直径为(220±6.33)μm,略大于于东祥等[19]报道的刺参卵径(140~170μm)。杂交子代的胚胎和幼体发育与中国刺参完全一致,但各个阶段的发育时间略有不同。在20~22℃时,中国刺参自交后代与杂交后代发育至初耳幼体的时间大致相同(24 h左右);前者经过第8天发育至大耳幼体,而杂交子代在第7天时即发育至大耳幼体;自交后代第10天时发育至樽形幼体,而杂交后代到第9天发育至樽形幼体。胡美燕等[15]的研究中也发现,相对于其他杂交或自交组合,日本红刺参♀×中国刺参♂的杂交子代在耳状幼体期间体长的生长最快,表现出一定程度的杂种优势(4.60%~6.90%),第8天生长率为67.8 μm/d,略高于本研究中第7天的生长率(59.38μm/d)。这表明,杂交苗在浮游幼体阶段发育速度较快,较于东祥等[18]报道的发育至樽形幼体时间提前1d左右。
在幼参培育阶段,当平均水温为25.2℃时,杂交苗的SGR略低于当地自交苗的SGR,但在夏季高温时期(水温27℃以上),杂交苗的生长明显优于自交苗。在此期间,自交苗活动能力减弱,摄食量明显减少,生长近于停滞(SGR为0.08%),而杂交苗的生长率随着水温的升高虽然也出现相应的下降(SGR为0.66%),但是下降幅度有限。这表明,在高温条件下,杂交苗较自交苗存在明显的生长优势。胡美燕和李琪[15]也发现了日本红刺参♀×中国刺参♂的杂交后代存在一定的生长优势,经过160 d的生长发育,这个组合的杂交后代在体长和体重增长方面优势非常明显,杂种优势率分别为8.39%~18.85%和23.21%~49.77%。本实验中发现幼参培育期间杂交苗的成活率略低于自交苗。王小兵等[20]的研究结果表明,单一的不同地理群体间的杂交所获得的子代,尤其对于杂交群体来自引进种群的杂交子代,虽然在生长速度上具有相当的杂种优势,但对于本地环境的适应能力较差。因而,在充分利用日本红刺参和中国刺参杂交子代生长优势的同时,应制定切实可行的选择育种方案提高杂交子代适应本地环境的能力。
本研究通过对中国刺参(♂)和日本红刺参(♀)杂交子代的胚胎发育和浮游幼体生长的观察,以及杂交和中国自交幼参生长和成活的比较,证实杂交与自交
后代幼体阶段的发育时序和形态特征基本一致,杂交苗发育至大耳幼体和樽形幼体的时间较自交苗短,在高温时期,杂交幼参存在明显的生长优势。此外,杂交刺参的体色美观,刺型优良,进一步开展杂交苗种的选择育种研究,有望培育出生长速度快、成活率高和抗逆能力强的刺参优良新品系。
[1] 中国渔业统计年鉴[M].北京:中国农业出版社,2009.
[2] Wang Z,Wang X,Zhang Q.Interfamilial hybridization between Paralichthys olivaceus and Kareius bicoloratus I.-Early development and growth characters[J].Journal of Fishery Sciences of China,2003,10(7):30-37.
[3] Purdom C E.Induced polyploidy in plaice(Pleuronectes platessa)and its hybrid with the flounder(Platichthys flesus)[J].Heredity,1972,29:11-23.
[4] 李思发,王成辉,刘志国,等.三种红鲤生长性状的杂种优势与遗传相关分析[J].水产学报,2006,30(2):175-180.
[5] 楼允东.鱼类育种学[M].北京:中国农业出版社,2001.
[6] Hedgecock D,McGoldrick D J,Bayne B L.Hybrid vigor in Pacific oysters:an experimental approach using crosses among inbred lines[J].Aquaculture,1995,137:285-298.
[7] Bayne B L,Hedgecock D,McGoldrick D,et al.Feeding behaviour and metabolic efficiency contribute to growth heterosis in Pacific oysters[Crassostrea gigas(Thunberg)][J].J Exp Mar Biol Ecol,1999,233:115-130.
[8] Launey S,Hedgecock D.High genetic load in the Pacific oyster Crassostrea gigas[J].Genetics,2001,159:255-265.
[9] 郑怀平,张国范,刘晓,等.不同壳色海湾扇贝(Argopecten irradians)家系的建立及生长发育研究[J].海洋与湖沼,2003,34(6):632-639.
[10] 刘小林,常亚青,相建海,等.栉孔扇贝不同种群杂交效果的研究[J].海洋学报,2005,27(2):135-140.
[11] 张国范,刘晓,阙华勇,等.贝类杂交及杂种优势理论和技术研究进展[J].海洋科学,2004,28(7):54-60.
[12] Kanno M,Kijima A.Genetic differentiation among three color variants of Japanese sea cucumber Stichopus japonicus[J].Fish Sci,2003,694:806-812.
[13] Kanno M,Suyama Y,Li Q,et al.Microsatellite analysis of Japanese sea cucumber,Stichopus(Apostichopus)japonicus,supports reproductive isolation in color variants[J].Mar Biotechnol,2006,8:672-685.
[14] Sun X J,Li Q,Kong L F.Comparative mitochondrial genomics within sea cucumber(Apostichopus japonicus):Provide newinsights into relationships among color variants[J].Aquaculture,2010,309:280-285.
[15] 胡美燕,李琪,孔令锋,等.中国刺参与日本红刺参杂交子一代的早期生长比较[J].中国海洋大学学报:自然科学版,2010,40(增刊2):375-380.
[16] Liu X L,Chang Y Q,Xiang J H.Hybridization effects of three populations of Chlamys Farreri from China,Korea,and Japan[J].J Shellfish Res,2004,23(2):501-505.
[17] 张国范,王继红,赵洪恩,等.皱纹盘鲍中国群体和日本群体的自交与杂交F1的RAPD标记[J].海洋与湖沼,2002,33(5):484-491.
[18] 王爱民,阎冰,叶力,等.马氏珠母贝不同地理种群内自繁和种群间杂交子一代主要性状的比较[J].水产学报,2003,27(3):200-206.
[19] 于东祥,孙慧玲,陈四清,等.海参健康养殖技术[M].北京:海洋出版社,2005.
[20] 王小兵,黄勃.杂色鲍杂交及选育育苗的效果研究[J].渔业现代化,2008,35(5):25-28.
Embryonic,Larval Development and Juvenile Growth in Hybrid Between Green(♂)and Red(♀)Sea Cucumber Apostichopus japonicus
SUN Xiu-Jun,LI Qi,KONG Ling-Feng,YU Rui-Hai
(The Key Laboratory of Mariculture,Ministry of Education,Ocean University of China,Qingdao 266003,China)
In present study,intraspecific hybridization was carried out between Green sea cucumber(♂)and Red sea cucumber(♀),Apostichopus japonicus.The embryonic development of hybrids was observed and the morphological characteristics of each period were described detailedly.Moreover,the growth rates and survival between hybrid juveniles and inbred Green juveniles were compared.At 20~22℃,the hybrid larvae required 24 h 38 min to develop to early auricularia,72 h 8 min to develop to mid-auricularia,and 167 h 8 min to develop to late auricularia.Furthermore,the hybrid larvae required 9 to develop to doliolaria.During the late stage of doliolaria,with the loss of cilia band,the larvae sank and became to settle on the bottom.From 38 to 51 after hatching,the average water temperature was 25.17℃,specific growth rate for weight(SGR)in the hybrid juveniles was 2.58%,while inbred juveniles grew at slightly higher SGR(3.03%).When the average water temperature rose to 27.17℃from 51 to 80 d,despite the growth rate of hybrid juveniles slowed down with SGR decreased by 0.66%,the average weight increased to(0.34±0.035)g at 80 d after hatching.However,with the decreased food intake of inbred juveniles,the SGR dropped sharply to a extremely low value(0.08%),with the average weight of(0.19±0.034)g at 80 d.There was no significant difference on survival between hybrid and inbred juveniles(82.2%and 73.9%).This indicated that the advantages of growth characteristics in the hybrid juveniles under the higher water temperature were obvious.
Apostichopus japonicus;hybridization;embryonic development;growth
S917.4
A
1672-5174(2012)04-026-07
山东省科技发展计划项目(2009GG10005013);高等学校博士学科点基金项目(20090132110018)资助
2011-04-09;
2011-08-18
孙秀俊(1981-),男,博士生,从事水产动物遗传育种学研究。E-mail:sxjrenp@hotmail.com
**通讯作者:E-mail:qili66@ouc.edu.cn
责任编辑 王 莉
