基因型和环境及其互作对糜子主要农艺性状的影响
2012-01-02李辛村杨天育张恩和
李辛村,杨天育,张恩和*
(1.甘肃农业大学农学院,甘肃 兰州730070;2.甘肃省农业科学院作物研究所,甘肃 兰州730070)
糜子(Panicummiliaceum)在我国具有悠久的栽培历史,是一种粮草兼用的作物,具有耐旱、耐瘠薄、适应性强、抗逆性强、适宜播期长、产量稳定的优良特性,在旱作农业生产中占有重要地位[1]。作物主要农艺性状不仅受其基因型的影响[2,3],而且也受环境及二者互作(genotype-environment interaction,G×E)的影响。G×E是产量稳定性的形成基础,也是决定基因型生态适应性的重要因子[4],G×E的值越大,则产量稳定性越差[5]。G×E决定了品种及其农艺性状在生产中的稳定性、适应性和差异性。施万喜[6]对旱地冬小麦(Triticumaestivum)新品种在不同生态条件下的丰产稳产性研究指出,环境和G×E对产量变化的影响远大于基因型。柴守玺等[7]对高寒农牧区不同试点和供水条件下小黑麦(Triticale)的研究表明,基因型间、环境间及G×E存在着极显著的差异,环境和G×E对产量的影响分别为基因型效应的25.9和2.1倍。李广昌[8]通过对不同生态条件下杂交早稻产量、生育期、有效穗、株高、穗长、每穗粒数、每穗实粒数、结实率、千粒重等9个产量相关性状的研究指出,环境和G×E对考察的9个性状影响较大。通过区域试验可鉴定品种的丰产性、适应性和稳定性,进而为农业生产提供具有推广价值的品种。
评价作物品种稳定性和适应性的方法很多,其中应用最为广泛的是AMMI模型(additive main effects and multiplicative interaction model),又称主效可加互作可乘模型[2,9]。该模型将方差分析与主成分分析相结合,不仅可分析基因型与环境的互作,还能对基因型相关性状的稳定性进行评价[10,11]。近年来,AMMI模型在小麦[6,12-14]、油菜(Brassicanapus)[15]、水稻(Oryzasativa)[8,16]、玉米(Zeamays)[17]等主要作物的产量和品质性状等方面得到广泛应用,同时还应用于区试中对地点鉴别力的判断[18],但在利用AMMI模型的分析中,大多采用产量和品质作为考察指标,而对区试糜子产量组成等主要农艺性状在AMMI模型中的表现则未见报道。北方地区是我国糜子的主要产区之一,研究糜子新品种的稳定性及其主要农艺性状的环境差异性,对于糜子新品种的推广应用具有重要意义。通过对糜子不同基因型主要农艺性状的均值比较,采用联合方差和AMMI模型分析基因型、环境及二者互作对糜子主要农艺性状的影响,以期选出适应性广、高产稳产的优良品种,并为糜子新品种的示范推广提供理论依据。
1 材料与方法
1.1 试验设计
试验于2008年在山西、内蒙古、陕西、宁夏、甘肃等地的11个试点进行,供试材料为国家第八轮区试的6个糜子品种:甘9109-6-1-1-2、甘9133-1-3-4-1、固02-25、固01-391、榆糜3号、伊8414-1-2-1,以榆糜3号为统一对照(CK)。各试点环境情况见表1。试验采用统一设计方案:完全随机区组,3次重复,小区面积10m2(2m×5m),行距33cm,人工开沟条播。各试点根据当地生产情况确定留苗密度(60~90万株/hm2)。各试点地力中等,田间管理略高于大田水平,田间记载和室内考种按统一方案执行,成熟后单独收获脱粒、称重计产。
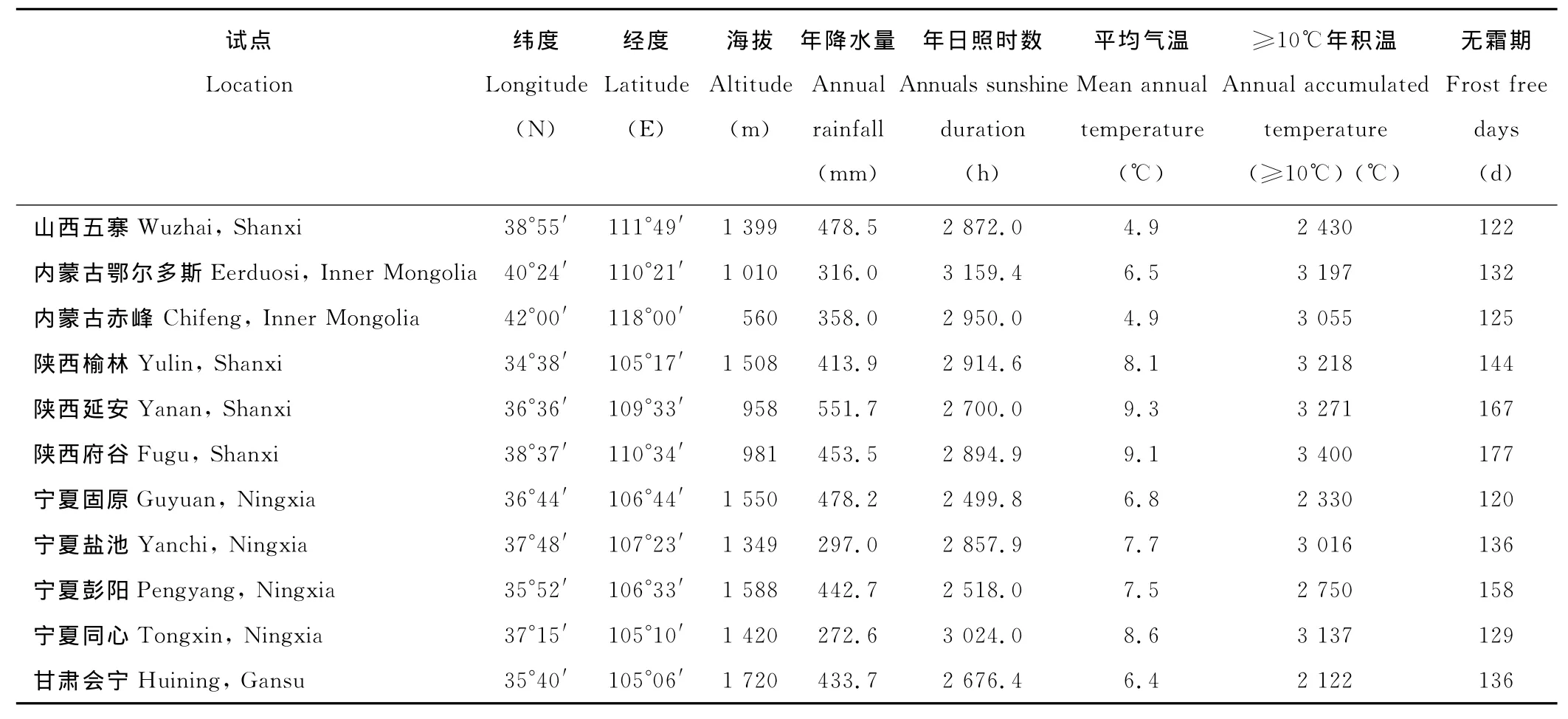
表1 试点环境因子Table 1 Environment factors of location
1.2 测定项目
测定不同试点不同基因型糜子的全生育期(从播种出苗至成熟的天数)、株高(从植株基部至穗顶端的长度)、主茎节数、穗粒重(每小区随机选取50穗混合脱粒,计算平均穗粒重)、千粒重(随机选取1 000粒种子,2次重复,取平均数)和产量(按小区面积实收籽粒重量计产折算)。
1.3 统计分析方法
首先进行联合方差分析,在基因型与环境互作效应显著的基础上,利用AMMI模型[11,14]进行品种稳定性分析,其数学模型[11,14]为:
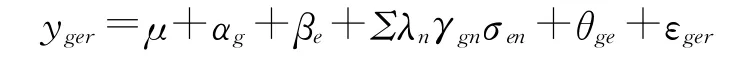
关于公式参数的解释详见参考文献[11,14],这里不再详述。
此研究取基因型在主成分效应达5%显著水平的IPCA空间中离原点的距离(欧氏距离Dg)[14]来度量基因型的相对稳定性,其值越小品种稳定性越高,对应的Dg值越大,则表示试点对品种差异的分辨力越强[14,19]。
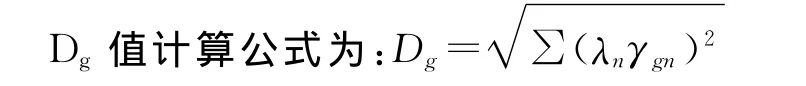
最大差异率(PR)计算公式:PR=(Max-Min)/Min×100%。式中,Max表示最大值,Min表示最小值。
数据处理与分析采用Microsoft Excel与唐启义和冯光明[20]的DPS 7.05数据处理系统。
2 结果与分析
2.1 基因型、环境及二者互作对糜子主要农艺性状的影响
生育期、株高、主茎节数、主穗长、穗粒重、千粒重、产量7个主要农艺性状的基因型和环境变异均达到1%的极显著水平(表2)。除株高、主穗长性状外,生育期、穗粒重、千粒重、产量4个主要农艺性状的基因型和环境的互作达到1%的极显著水平,主茎节数的基因型和环境互作达到5%的显著水平。从基因型、环境及二者互作所占总平方和的百分比可以看出,各性状的基因型变异均小于二者互作变异和环境变异。对于产量性状,基因型、环境和互作均达极显著水平。其中,环境变异的平方和占整个处理平方和的86.6%,而基因型和互作变异的平方和分别占2.0%和11.5%。表明处理变异平方和绝大部分归因于环境间的差异,而二者互作的变异又大于基因型变异。基因型对各性状的作用大小依次为主茎节数>千粒重>株高>主穗长>穗粒重>生育期>产量;环境的作用大小依次为株高>产量>主穗长>千粒重>生育期>主茎节数>穗粒重;二者互作的作用大小依次为穗粒重>生育期>主茎节数>主穗长>产量>千粒重>株高。说明基因型对主茎节数、环境对株高、二者互作对穗粒重的影响较大。
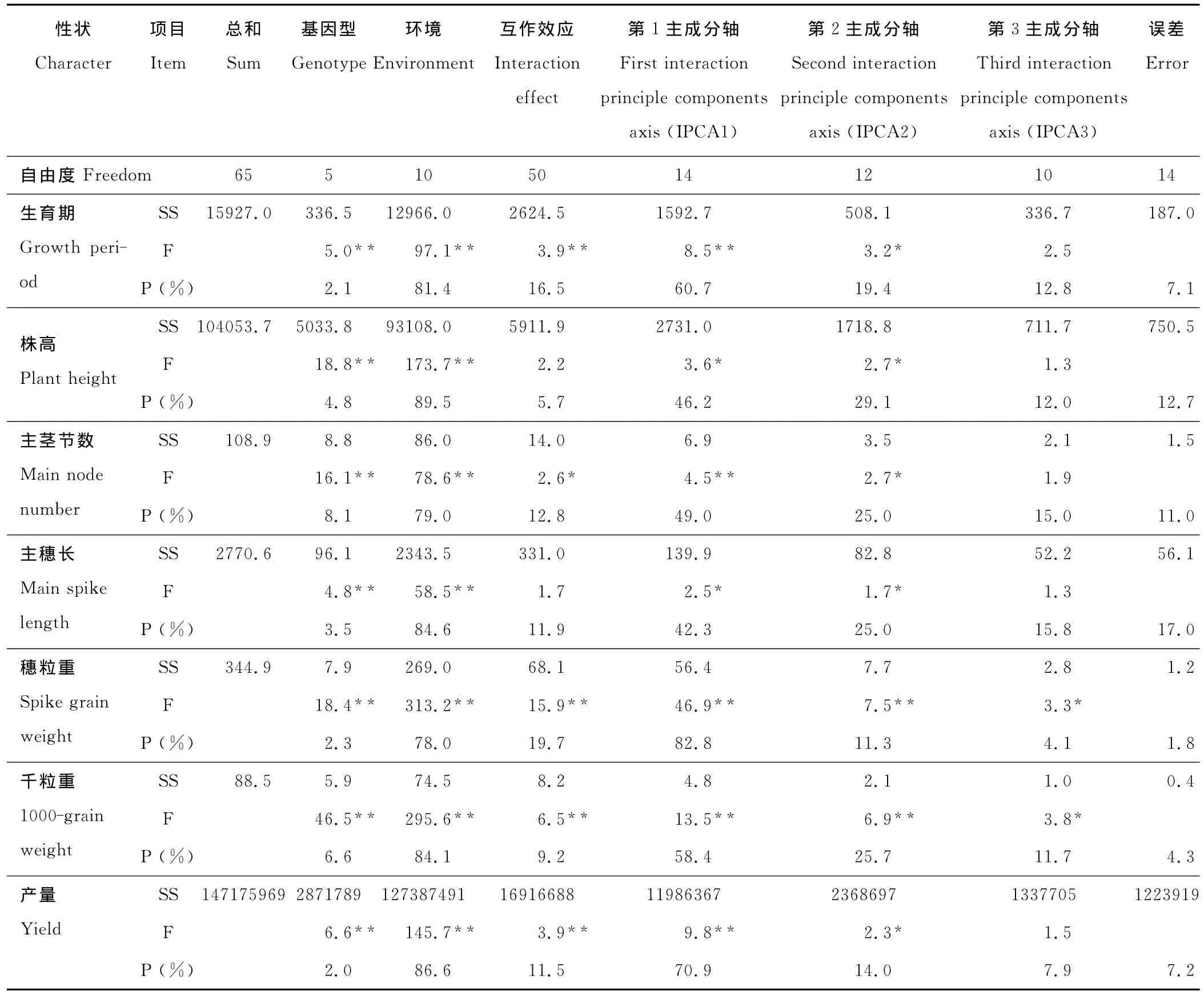
表2 基因型与环境互作的方差分析和AMMI模型分析结果Table 2 The analysis results between genotype environment interaction variance analysis and AMMI model on P.miliaceum
2.2 不同基因型糜子主要农艺性状的环境变异
同一基因型糜子在不同环境下各性状存在明显差异(表3)。各参试品种7个农艺性状指标的变异系数均值中,产量最大,达42.72%,其次是穗粒重和株高,分别为37.90%和26.74%,生育期变异系数最小,为14.89%。
基因型间各性状均值,株高榆糜3号(163.1cm)最高,固01-391(141.3cm)最低;各参试品种均值低于对照。主茎节数甘9109-6-1-1-2(8.9节)最多,固01-391(7.8节)最少;甘9109-6-1-1-2和甘9133-1-3-4-1高于对照,固01-391和伊8414-1-2-1低于对照。主穗长榆糜3号(33.7cm)最大,固01-391(29.8cm)最小;各品种均值低于对照。穗粒重伊8414-1-2-1(6.8g)最大,固02-25(5.7g)最小;除固02-25外,其他品种均值高于对照。千粒重榆糜3号(7.7g)最大,固01-391(6.8g)最小;各品种均值低于对照。产量甘9133-1-3-4-1(3 802.7kg/hm2)最高,固02-25(3 219.6kg/hm2)最低;除固02-25外,各品种产量明显高于对照。

表3 不同基因型糜子主要农艺性状的平均值和变异系数比较Table 3 Effect of P.miliaceumgenotype on average value and variation coefficient of main agronomic traits
基因型间各性状变异系数,生育期变异程度伊8414-1-2-1(15.91%)最大,甘9109-6-1-1-2(12.71%)最小;株高变异 程 度 固 01-391(30.83%)最 大,甘 9109-6-1-1-2(23.31%)最 小;主 茎 节 数 变 异 程 度 甘 9109-6-1-1-2(17.25%)最大,甘 9133-1-3-4-1(11.46%)最 小;主 穗 长 变 异 程 度 固 01-391(28.35%)最 大,伊 8414-1-2-1(17.95%)最小;穗粒重变异程度甘9109-6-1-1-2(40.19%)最大,伊8414-1-2-1(34.60%)最小;千粒重变异程度甘9109-6-1-1-2(18.23%)最大,甘9133-1-3-4-1(10.99%)最小;产量变异程度伊8414-1-2-1(45.40%)最大,固02-25(36.52%)最小。基因型变异程度小,表明品种在各环境中静态稳定性好,但不利于高产栽培,一般变异程度小而均值高的品种比较好。
基因型间各性状最大差异率(PR)变化趋势与对应的变异系数(CVg)一致,但最大差异率显著高于对应变异系数,说明最大差异率对环境变异敏感,更能够如实反映各农艺性状对环境变异的极端差异。各性状变异系数和最大差异率均值产量最大,生育期最小,说明环境对产量的影响较大,对生育期的影响较小。
2.3 不同环境间糜子主要农艺性状的差异
各试点平均生育期宁夏同心最长(表4),陕西府谷最短,相差47d;平均株高陕西榆林最高,比最低的宁夏固原高125.7cm;平均主茎节数最多的是陕西府谷,比最少的宁夏固原多3.0节;平均主穗长以内蒙古赤峰最长,宁夏固原最短,相差20.3cm;平均穗粒重陕西延安最高,宁夏同心最低,相差7.1g;平均千粒重以内蒙古鄂尔多斯最高,宁夏盐池最低,相差3.9g;各试点平均单产陕西延安最高,宁夏彭阳次之,宁夏固原最低,表明各试点间的环境条件有较大差异。

表4 不同环境间糜子主要农艺性状差异比较Table 4 Effect of environment on main agronomic traits of P.miliaceum
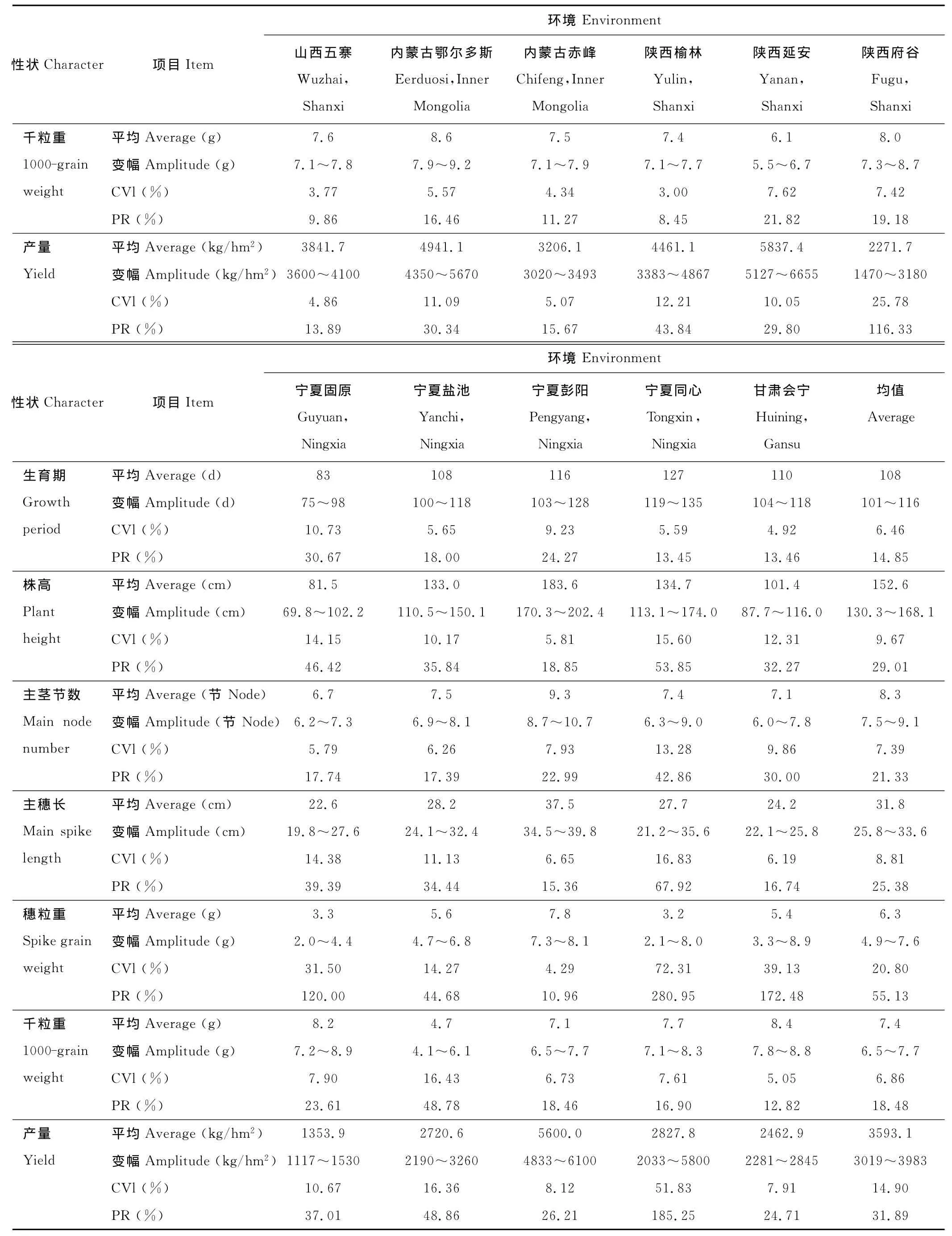
续表4 Continued
环境间各性状变异系数,生育期宁夏固原变异程度最大,陕西榆林最小;株高宁夏同心变异程度最大,陕西榆林最小;主茎节数和主穗长宁夏同心变异程度最大,内蒙古鄂尔多斯最小;穗粒重宁夏同心变异程度最大,宁夏彭阳最小;千粒重宁夏盐池变异程度最大,陕西榆林最小;产量性状变异程度宁夏同心最大,山西五寨最小。结果表明,宁夏彭阳和陕西延安试点产量较高,变异程度较小,对品种鉴别力较好。
环境间各性状最大差异率变化趋势与对应的变异系数一致,但其值与对应变异系数间的差异相对减小。各性状变异系数和最大差异率均值穗粒重最大,生育期最小,说明穗粒重受基因型的影响较大,生育期受基因型的影响较小。
2.4 不同基因型糜子主要农艺性状品种稳定性分析
不同基因型糜子生育期、株高、主茎节数、主穗长、穗粒重、千粒重和产量性状5%显著水平的主成分效应之和(IPCA1+IPCA2),分别解释了相应基因型与环境互作变异平方和的80.1%,75.3%,74.0%,67.3%,94.1%,84.1%和84.9%(表2)。因此,利用IPCA1和IPCA2计算稳定性参数Dg值(表5),能较准确地判断糜子基因型主要性状的品种稳定性。生育期和产量稳定性最好的基因型是榆糜3号(CK)(表5);株高和主茎节数稳定性最好的基因型是固02-25;主穗长稳定性最好的基因型是甘9109-6-1-1-2;穗粒重和千粒重稳定性最高的基因型是固01-391。结果表明,对照基因型榆糜3号的产量和生育期稳定性高于其他基因型,说明其适应性强。
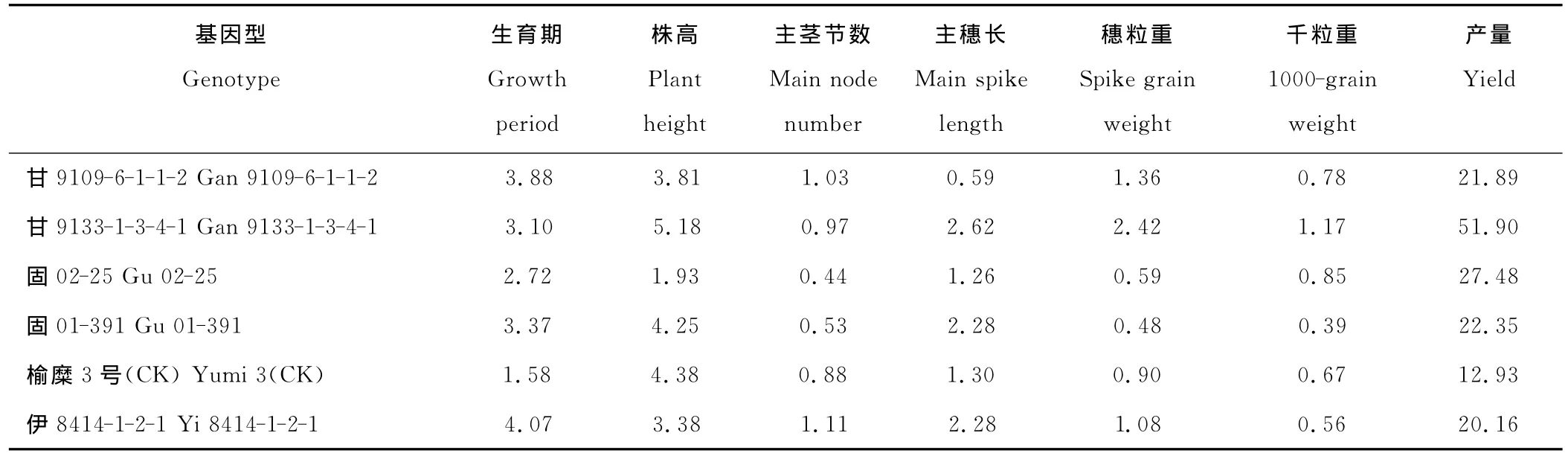
表5 不同基因型糜子主要农艺性状的稳定性参数(Dg)Table 5 The stability of different P.miliaceumagronomic traits(Dg)
3 讨论
作物品种区试中基因型和环境互作现象普遍存在。由于作物农艺性状的表现是在基因型与环境共同作用下形成的,环境条件的不同可能导致基因表达方式或程度的差异。有关研究表明,环境间、基因型与环境互作引起的变异远大于基因型间的变异。Gauch和Zobel[21]研究指出,在影响作物产量等主要农艺性状变异因素中,来自基因型间、环境间和二者互作的比例大致为10%,70% 和20%,在互作中,地点×年份>基因型×地点×年份>基因型×地点>基因型×年份。Rosella等[22]曾在地中海不同环境下对春小黑麦稳定性研究发现,基因型×环境引起的单位面积籽粒产量差异远大于基因型间本身的遗传差异,其中环境、基因型、基因型与环境互作对籽粒产量变异的贡献分别是85%,3%和11%。Romagosa和Fox[23]在总结了100多个环境产量试验资料后,甚至得出来自环境变异的比例高达80%~90%。李广昌[8]研究表明,不同品种水稻的穗粒数和产量受环境及基因型×环境影响较大,千粒重、穗长和有效穗受环境及基因型×环境影响较小。本研究表明,环境及其基因型与环境互作是引起糜子产量、穗粒重等主要农艺性状差异的主要原因,也就是说,环境及其基因型与环境互作对糜子主要农艺性状有显著的影响,这一结果与常磊等[24]、Roozeboom等[25]在小麦等作物上的研究结果相一致。由于基因与环境的互作远大于基因型效应,且有些糜子品种对环境具有特殊适应性。因此,在适应当地条件扩展生态育种的同时,应重视品种布局,将品种种植在最适合的环境条件下,以充分发挥基因与环境的正向互作效应。
作物的基因型与环境互作决定了作物品种在生产中的稳定性和适应性。张锡顺等[15]用线性回归模型分析油菜区试数据时,剩余残差占互作的74.6%,而在AMMI模型中,残差占互作的16.6%。说明使用AMMI模型能提高估计的准确性。本研究表明,残差占互作的比例较小,分别为7.1%(生育期)、1.5%(株高)、11.0%(主茎节数)、17.0%(主穗长)、1.8%(穗粒重)、4.3%(千粒重)和7.2%(产量)。主要是由于 AMMI分析中,显著的主成分轴(IPCA1和IPCA2)用尽量小的自由度捕捉尽量大的平方和,而剩余大自由度对应小的平方和,作为试验误差处理,可提高试验的精确度。当显著的IPCA1轴多于2个以上时,可通过欧氏距离(D值)大小直观得出各参试品种的稳定性和较适应地区。
基于欧氏距离得到的AMMI模型稳定性参数Dg,能够全面利用显著或极显著乘积项的信息,是定量判别品系和地点稳定性的重要参考指标[2]。本研究利用稳定性参数Dg值考察了不同基因型糜子品种在各主要农艺性状中的稳定性表现,表明农艺性状在基因型的稳定性表现存在差异。不同品种糜子7个主要农艺性状中,产量的稳定性随基因型不同而变化较大(12.93≤Dg≤51.9),榆糜3号(Dg≤12.93)的产量稳定性最高,其次是甘9109-6-1-1-2(Dg=21.89)和固01-391(Dg=22.35),甘9133-1-3-4-1(Dg=51.9)的稳定性较差。
通过比较各个参试地点的鉴别力和品种的稳产性,可以对试验点和品种做出评判,有利于区域试验地点的取舍,提高试验的准确性和效率。本研究表明,糜子主要农艺性状在不同基因型以及环境之间存在极显著差异:基因型对主茎节数,环境对株高,基因型与环境互作对穗粒重的影响较大;各品种农艺性状变异系数最大的是产量,最小的是生育期;各试点产量最高的是甘9133-1-3-4-1,品种稳定性最好的是榆糜3号;宁夏彭阳和陕西延安试点对品种鉴别力较好。本研究所用数据为1年,关于这些品种的稳产性、丰产性和适应性有待通过多年数据进一步验证,以比较结果的重复性。
[1]董孔军,何继红,杨天育.甘肃省糜子区试品种(系)稳定性的AMMI分析[J].作物杂志,2008,(3):73-76.
[2]张泽,鲁成,向仲怀.基于AMMI模型的品种稳定性分析[J].作物学报,1998,24(3):304-309.
[3]蔺海明,周建军,王蒂,等.大整薯稀播对马铃薯农艺性状和产量的影响[J].草业学报,2011,20(3):304-308.
[4]衣兰智,李长忠,刘洪庆,等.不同苜蓿品种在青岛地区的适应性[J].草业学报,2011,20(2):147-155.
[5]Santiveri F,Royo C,Romagosa I.Patterns of grain filling of springand winter hexaploid triticales[J].European Journal of Agronomy,2002,16:219-230.
[6]施万喜.利用AMMI模型分析陇东旱地冬小麦新品种(系)丰产稳产性[J].干旱地区农业研究,2009,27(3):37-43.
[7]柴守玺,常磊,杨蕊菊,等.小黑麦基因型与环境互作效应及产量稳定性分析[J].核农学报,2011,25(1):155-161.
[8]李广昌.福建省龙岩市杂交早稻产量相关性状的基因型与环境变异[J].中国农学通报,2009,25(9):58-64.
[9]莫惠栋,曹桂英.作物品种区试资料的非参数度量[J].中国农业科学,1999,32(4):85-91.
[10]穆培源,庄丽,张吉贞,等.作物品种稳定性分析方法的研究进展[J].新疆农业科学,2003,40(3):142-144.
[11]郑轶琦,臧国长,郭海林,等.假俭草杂交后代生殖性状遗传及相关性分析[J].草业学报,2011,20(2):283-289.
[12]常磊,柴守玺.AMMI模型在旱地春小麦稳定性分析中的应用[J].生态学报,2006,26(11):3677-3684.
[13]郭世华,王洪刚.基因型与环境及其互作对我国冬小麦部分品质性状的影响[J].麦类作物学报,2006,26(1):45-51.
[14]任勇,李生荣,杜小英,等.不同基因型小麦新品系主要性状稳定性分析[J].中国农学通报,2007,23(4):154-158.
[15]张锡顺,杨德,谢永俊,等.AMMI模型应用于油菜区域试验的分析[J].西南农业学报,2001,14(2):27-30.
[16]刘文江,李浩杰,汪旭东,等.用AMMI模型分析杂交水稻基本性状的稳定性[J].作物学报,2002,28(4):569-573.
[17]吴渝生,李本狲,顾红波,等.甜玉米品种稳定性的AMMI模型分析[J].华中农业大学学报,2003,22(1):4-8.
[18]李本贵,阎俊,何中虎,等.用AMMI模型分析作物区域试验中的地点鉴别力[J].作物学报,2004,30(6):593-596.
[19]Li B G,Yan J,He Z H,etal.Analyzing site discrimination in crop regional yield trials by AMMI model[J].Acta Agronomica Sinica,2004,30(6):593-596.
[20]唐启义,冯光明.实用统计分析及其DPS数据处理系统[M].北京:科学技术出版社,2002:262-267.
[21]Gauch H G,Zobel R W.AMMI analysis of yield trails[A].In:Kang M S,Gauch G H.Genotype-by-Environment Interaction[M].Boca Raton Florida:CRC Press,1996:85-122.
[22]Rosella M,Francesco G,Mauro D.Factors affecting the genotype×environment interaction in spring triticale grown in a Mediterranean environment[J].Euphytica,2001,121:317-324.
[23]Romagosa I,Fox P N.Genotype×environment interaction and adaptation[A].In:Hayward M D,Bosemark N O,Romagosa I.Plant Breeding:Principle and Prospects[M].London:Chapman and Hall,1993:373-390.
[24]常磊,柴守玺,杨德龙,等.我国旱地春小麦产量及主要农艺指标的变异分析[J].应用生态学报,2010,21(11):2821-2829.
[25]Roozeboom K L,Schapaugh W T,Tuinstra M R,etal.Testing wheat in variable environments:Genotype,environment,interaction effects,and grouping test locations[J].Crop Science,2008,48:317-330.
