水土保持林生长过程及碳密度的动态变化
2012-01-02张建军徐佳佳李慧敏
张建军,徐佳佳,李慧敏
(北京林业大学水土保持学院,100083,北京)
目前,气候变暖已成为全球关注的热点,CO2等温室气体排放量的增加是导致气候变暖的主要原因[1]。森林生态系统在调节全球碳平衡、减缓大气中CO2等温室气体浓度上升以及维护全球气候等方面具有不可替代的作用[2-3];因此,开展森林植被吸收和固定CO2能力的定量评价研究已成为热点和前沿[4]。
刺槐(Robinia pseudoacacia L.)是蝶形花科刺槐属的乔木,有较强的适应性和抗逆性,是我国主要的水土保持造林树种,更是黄土高原水土保持速生树种的典型代表。在黄土高原地区营造有大面积的刺槐人工林[5],这些刺槐人工林改善了当地的生态环境,对治理黄土高原水土流失发挥了积极作用;但人工刺槐林根系分布广、耗水量较大[6],造林地深层土壤干燥化现象普遍发生,形成了大面积低效刺槐林[7],成为黄土高原人工植被建设的重大生态隐患[8]:因此,正确评价黄土高原刺槐人工林的生态服务功能是目前亟待解决的问题。
水土保持林作为水土流失地区特殊的人工生态系统,通过光合作用将CO2转化成有机物后固定在植物体内,这些有机物除去自身的呼吸消耗、枯落物分解后,以生物量的形式保存在植物体内;因此,水土保持林作为水土流失地区特有的植被类型,不仅起到了防治水土流失的作用,在碳汇方面的功能也应该受到重视。国内学者围绕森林生态系统的碳密度、碳储量做了大量的研究工作,并取得了显著成就[9-11];但有关森林生态系统碳密度、碳储量的研究多注重天然林,对人工林碳密度、碳储量的研究相对较少[12],而对黄土区水土保持林碳密度、碳储量的研究更为少见。为此,笔者以黄土高原主要的水土保持林——刺槐林为研究对象,在探讨其生长规律的基础上,研究其生物量和碳密度的动态变化,以期为水土保持植被生态服务功能评价提供参考。
1 研究区概况
研究区位于山西省吉县红旗林场的马莲滩,地理坐标E110°45'04″~110°47'38″,N36°00'15″~36°04'32″,属于晋西黄土残塬沟壑区,海拔965 ~1 345 m。年平均降水量575.9 mm,年平均水面蒸发量1 732.9 mm,年平均气温10 ℃。土壤为褐土,黄土母质。森林覆盖率为45%,主要造林树种有刺槐和油松(Pinus tabulaeformis),灌木树种主要有沙棘(Hippophae rhamnoides)和虎榛子(Ostryopsis davidiana)等。
2 研究方法
在对研究区刺槐人工林调查的基础上,选择不同密度的标准地进行样方调查,样方面积为400 m2。调查样方基本情况见表1。

表1 样地基本情况Tab.1 Basic condition of sample plot
在样方内进行每木检尺,测量刺槐的树高、胸径。根据测定的树高、胸径数据计算出标准地的平均树高和平均胸径,以此为依据选择3 株标准木。
将标准木锯倒后摘下所有叶片称量,作为该标准木的鲜叶生物量,在摘下的叶子中抽取500 g 作为样品装入保鲜袋,带回实验室进行含水率的测定;将伐倒标准木所有的枝条锯下后称量,作为枝条生物量的鲜质量,并取样500 g 带回实验室进行含水率的测定;将去除枝条的树干称量作为树干生物量鲜质量,并将树干以1 m 为区分段截取树干解析用的圆盘,带回实验室进行树干解析,同时对每个圆盘进行称量。
采用全根挖掘法测定标准木地下部分所有根系的鲜质量,并取样1 000 g 装入保鲜袋,带回实验室用于测定根系含水率。
在室内对圆盘进行分析后,与树叶样品、枝条样品、根系样品一起放入80°的烘箱内烘干至恒质量后测定含水率。
利用树干、枝条、根系、叶的含水率,分别计算出标准木的树干、枝条、根系、叶的干质量生物量后,将各部分相加得出标准木的总生物量。根据马钦彦等[13]测定的阔叶树树干、枝条、根系、树叶的含碳率计算出刺槐林的碳储量。
3 结果与分析
3.1 树高生长分析
表2 为刺槐林树高生长分析表。可以看出,树龄4 a 的刺槐林树高总生长量阳坡比阴坡高0.24 m,而5 a 以上的刺槐林树高总生长量均为阴坡大于阳坡。这主要是由于树龄小于4 a 时,刺槐林尚未郁闭,土壤中的水分尚能满足刺槐高生长之需。当刺槐的树龄达到5 a 后,林木耗水不断增加,阳坡林地的土壤水分与林木耗水的供需矛盾开始加剧,从而导致阳坡刺槐林的高生长较阴坡差。
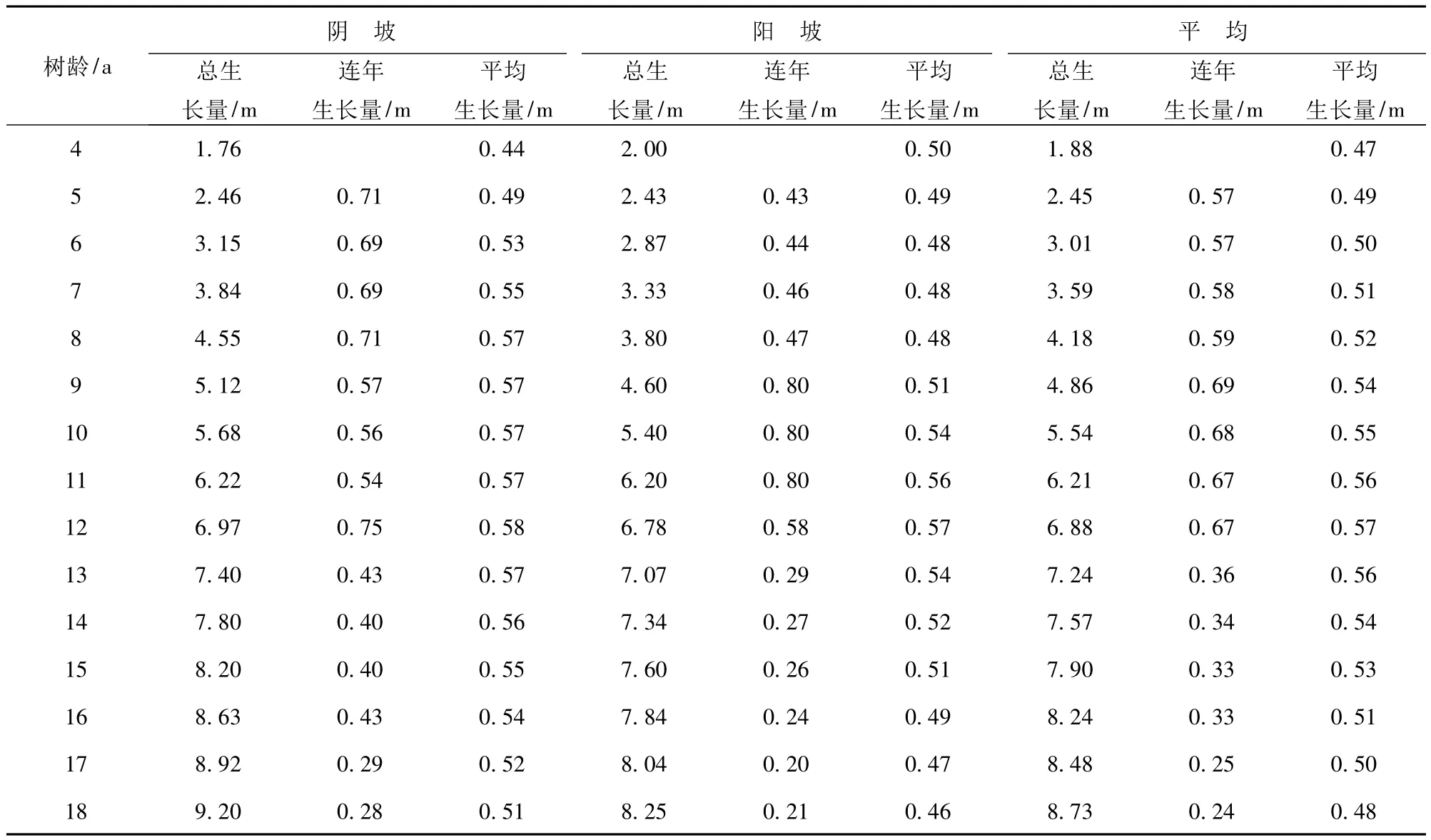
表2 刺槐林树高生长分析表Tab.2 Analysis of the tree height growth of Robinia pseudoacacia
图1 为刺槐树高的总生长量、连年生长量、平均生长量随树龄的变化图。可见,树龄18 a 的刺槐平均树高可达8.7 m。从树高总生长量变化曲线可见,以12 a 树龄为界,刺槐高生长可分为2 个阶段,第1 阶段的树龄小于12 a,树高生长属于快速增长阶段,树高的增高速率可达0.63 m/a,树龄12 a 时,树高可达6.9 m,当树龄超过12 a 后,树高生长速率变慢,进入缓慢增长的第2 阶段,树高增高的速率只有0.32 m/a,到树龄18 a 时,树高才到8.7 m,可见,树高增高速率第2 阶段明显缓于第1 阶段,第2 阶段只有第1 阶段的50.8%。
从连年生长量曲线和平均生长量曲线也可以看出,树龄达到12 a 后,树高的连年生长量曲线与平均生长量曲线相交,即在树龄小于12 a 时,树高的连年生长量明显大于平均生长量,而当树龄大于12 a 后,树高的连年生长量显著小于平均生长量。可见,当刺槐林龄达到12 a 后,刺槐高生长开始进入成龄高度,此后高生长逐渐缓慢。

图1 刺槐树高生长过程Fig.1 Height growth process of Robinia pseudoacacia
利用理查德生长方程拟合的刺槐高生长模型为

式中:H 为树高,m;a 为树龄,a。
利用该模型可以计算出当刺槐树龄达到20、30和40 a 时,树高分别可以达到12.4、14.3 和15.1 m。树龄在21 ~30 a 期间,树高的平均生长率只有0.16 m/a,树龄在31 ~40 a 期间,树高的平均生长率只有0.07 m/a。树龄19 ~40 a 的刺槐平均生长率为0.13 m/a,而树龄18 a 之前树高增高速率为0.32 m/a。可见,树龄18 a 以后,刺槐的高生长趋于缓慢。
3.2 胸径生长分析
表3 为刺槐林胸径生长分析表。可以看出:树龄7 a 的刺槐胸径总生长量阴坡为2.46 cm,阳坡为2.44 cm,二者相差无几;但当树龄小于7 a 时,刺槐胸径总生长量阳坡均比阴坡大。可能的原因是:当刺槐树龄较小时,土壤水分并不是胸径生长的限制因素,或胸径生长与土壤水分供应的矛盾不突出;当树龄大于7 a 后,刺槐胸径总生长量阴坡大于阳坡,这是由于阳坡太阳辐射强烈,土壤供水能力与刺槐耗水能力矛盾加剧所致。

表3 刺槐林胸径生长分析表Tab.3 Analysis of DBH growth of Robinia pseudoacacia
图2 为刺槐胸径总生长量、连年生长量、平均生长量随树龄的变化图。可以看出,树龄18 a 的刺槐胸径总生长量可达7.14 cm。从刺槐胸径总生长量变化曲线可见,以12 a 的树龄为界,刺槐胸径总生长量也可分为2 个阶段,第1 阶段的树龄小于12 a,胸径生长属于快速增长阶段,胸径的增长速率为0.53 cm/a,到树龄为12 a 时,刺槐胸径总生长量可以达到5.19 cm;第2 阶段的树龄为12 ~18 a,胸径生长速率变缓,胸径增长速率只有0.32 cm/a,属于缓慢增长阶段,到树龄18 a 时,刺槐胸径总生长量达到了7.14 cm。可见,刺槐胸径总生长量增大的速率第2 阶段明显缓于第1 阶段,第2 阶段只有第1阶段的60.38%。
从胸径连年生长量曲线和平均生长量曲线也可以看出,树龄达到12 a 后,胸径连年生长量曲线与平均生长量曲线相交,即在树龄小于12 a 时,刺槐胸径连年生长量明显大于平均生长量,而当树龄大于12 a 后,刺槐胸径连年生长量显著小于平均生长量。可见,树龄达到12 a 时刺槐胸径的生长基本达到成龄阶段,此后胸径生长趋于缓慢。
利用理查德生长方程拟合出的刺槐胸径的生长模型为

式中D 为胸径,cm。
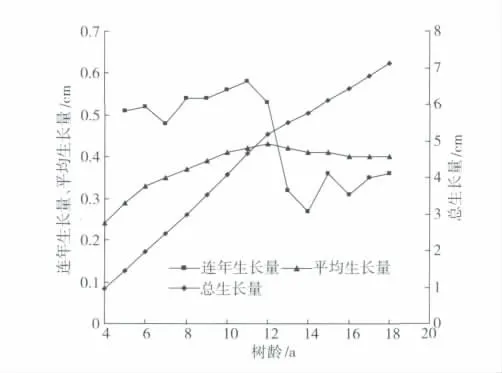
图2 刺槐胸径生长过程Fig.2 DBH height growth process of Robinia pseudoacacia
利用该模型可以计算出当树龄达到20、30 和40 a 时,刺槐胸径的总生长量分别可以达到7.8、10.2 和11.5 cm。当刺槐树龄在21 ~30 a 期间时,胸径的平均生长率只有0.21 cm/a,树龄在31 ~40 a期间时,胸径的平均生长率只有0.12 cm/a。利用胸径生长模型计算出刺槐林树龄在19 ~40 a 期间平均生长率只有0.18 cm/a,而在树龄18 a 之前,其胸径增长速率为0.32 cm/a,可见,当树龄大于18 a后,刺槐的胸径生长速率明显变慢。
3.3 胸径与生物量的关系
水土保持林调查中,胸径的测定相对容易,且测量精度较高,而生物量通常需要将对象木伐倒进行测量,属于破坏性调查,为此建立胸径与生物量的关系具有重要意义。对研究区42 株不同胸径刺槐的生物量进行测定,利用其中35 株的数据拟合出了刺槐单株生物量(干质量)(y)与胸径(x)的关系(图3)。可以看出,刺槐单株生物量(干质量)与胸径成幂函数关系。利用剩余7 株的数据对刺槐单株生物量(干质量)与胸径的关系进行验证,结果见表4。

图3 刺槐单株生物量与胸径的关系Fig.3 Relationship between biomass and DBH of a single Robinia pseudoacacia
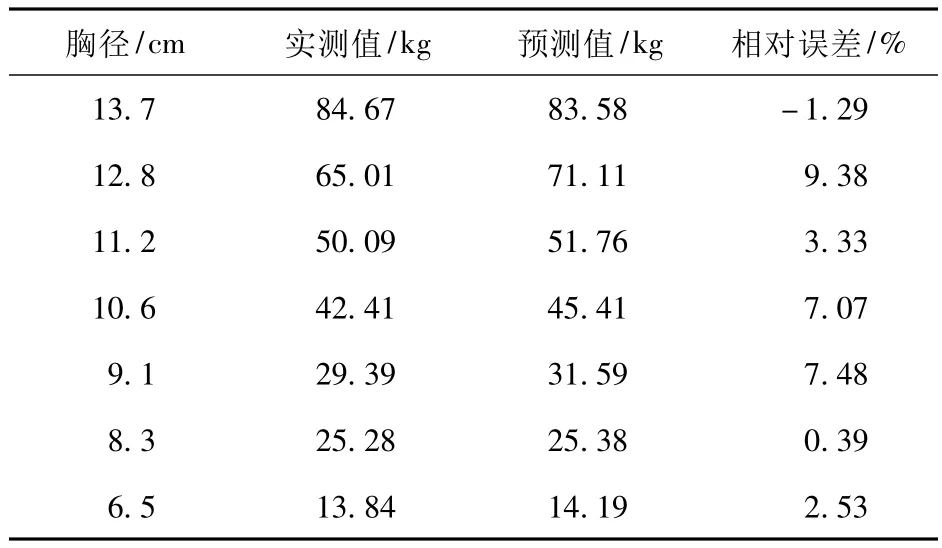
表4 刺槐单株生物量预测值与实测值的对比Tab.4 Biomass comparison between the predictive value and measured value of single Robinia pseudoacacia plant
刺槐单株总生物量、叶生物量、根生物量与胸径的关系式如下:

式中:W总为刺槐单株总生物量,kg;W叶为刺槐单株叶生物量,kg;W根为刺槐单株根生物量,kg。
4 水土保持林分生物量与碳密度的动态变化关系
研究区刺槐水土保持林是山西吉县红旗林场于20 世纪70 年代营造的,平均密度为1 500 株/hm2,平均胸径为7.8 cm。利用胸径与单株生物量的关系可以计算出研究区刺槐水土保持林目前的单位面积生物量约为32.84 t/hm2。
碳密度是单位面积生物量与含碳率的乘积,据王效科等[14]的研究,含碳率在不同植物间变化不大,因此,为简便起见,采用马钦彦等[13]对华北主要森林类型建群种含碳率的研究结果,选取华北地区落叶阔叶树种含碳率的平均值0.488 作为刺槐水土保持林的含碳率,计算出目前研究区刺槐水土保持林的碳密度为16.03 t/hm2。
利用前面得出的刺槐胸径生长模型D=12.984(1-e-0.066a)1.652和胸径与生物量的关系W总=0.165 4D2.3784,对研究区刺槐胸径、生物量、碳密度的动态变化进行计算和预测,结果见图4 和表5。
从图4 可见,研究区刺槐生物量的连年生长量在树龄21 a 时达到最大,为3.11 t/hm2,树龄21 a 后生物量的连年生长量开始下降,即从树龄21 a 开始,刺槐林生物量的增长速率逐渐下降,到树龄35 a时生物量的连年生长量与平均生长量相交,此时刺槐水土保持林已经达到成熟林,总生物量可以达到72.79 t/hm2,碳密度为35.52 t/hm2(表5)。
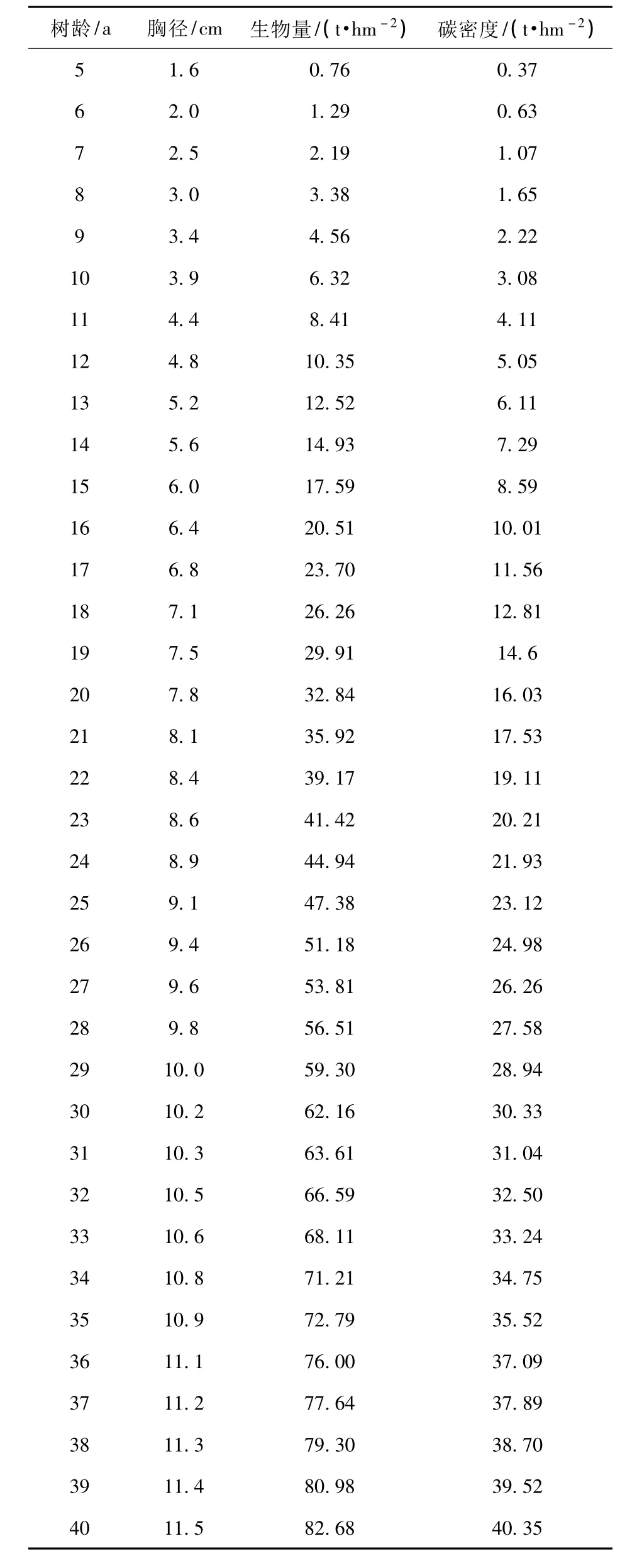
表5 刺槐水土保持林胸径、生物量、碳密度的动态变化Tab.5 Dynamic changes of DBH,biomass,carbon intensity of Robinia pseudoacacia forests for soil and water conservation
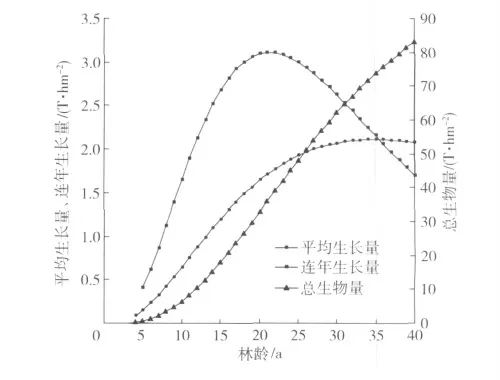
图4 刺槐水土保持林生物量变化图Fig.4 Biomass changes of Robinia pseudoacacia forests for soil and water conservation
5 结论与讨论
1)研究区刺槐水土保持林的高生长可分为2个阶段,树龄12 a 以前为快速生长阶段,树高的生长速率可达0.63 m/a,树龄12 ~18 a 为缓慢生长阶段,树高的生长速率只有0.32 m/a。利用树高生长模型预测得出树龄19 a 以后树高的生长速率只有0.13 m/a。刺槐林龄达到12 a 后进入成林高度,高生长逐渐缓慢。
2)以树龄12 a 为界,刺槐水土保持林的胸径生长分为2 个阶段,树龄12 a 以前胸径平均生长率为0.53 cm/a,2 ~18 a 胸径平均生长率为0.32 cm/a。利用胸径生长模型预测得出树龄19 a 以后胸径的生长速率只有0.18 cm/a。刺槐树林达到12 a 后胸径生长逐渐变慢。
3)刺槐的总生物量、叶生物量、根生物量均与胸径成幂函数关系。通过模拟计算得出,树龄达到21 a 时,刺槐水土保持林生物量的连年生长量最大,达3.11 t/hm2,此后生物量的增长速率逐渐下降,到树龄35 a 时,生物量的连年生长量与平均生长量相交,成为成熟林,总生物量达到72.79 t/hm2,碳密度为35.52 t/hm2。
调查地区刺槐水土保持林目前的碳密度仅为12.81 t/hm2,远低于刘国华等[15]提出的我国森林平均碳密度38.7 t/hm2,而当刺槐水土保持林达到成熟林的35 a 后,其他密度可以达到35.52 t/hm2,与我国森林的平均碳密度已非常接近,这说明生长在水土流失地区的水土保持林,虽然生存条件苛刻,但这些水土保持林不仅能够起到防治水土流失、保护土壤资源、改善当地生态环境的生态服务功能,同时具有较强的碳汇能力,为固碳减排起到了积极作用;因此,在评价水土保持林生态服务功能时,必须充分考虑水土保持林的固碳效益。
刺槐水土保持林达到树龄35 a 后,生物量的连年增长量小于平均增长量,此后其固定CO2的能力逐年下降;因此,应该对这种成熟林进行更新改造,以防止水土保持林碳汇能力的衰退和降低。
[1] Caldeira K,Duffy P B.The role of the southern ocean in uptake and storage of anthropogenic carbon dioxide[J].Science,2000,287:620-622
[2] Banaitis M R,Langley-Tumbaugh S J,Aboueissa A A.Variations of soil organic carbon in three urban parks a maine case study[J].International Journal of Applied Environmental Sciences,2007,2:119-128
[3] Paul K I,Polglase P J,Nyakuengama J G,et al.Change in soil carbon following afforestation[J].Forest Ecology and Management,2002,168:241-257
[4] Fang Jingyun,Chen Anping,Peng Changhui,et al.Changes in forest biomass carbon storage in China between 1949 and 1998[J].Science,2001,292:2320-2322
[5] 邓磊,张文辉.黄土沟壑区刺槐人工林的天然发育规律[J].林业科学,2010,46(12):15-21
[6] 张建军,张岩,张波.晋西黄土区水土保持林地的土壤水分[J].林业科学,2009,45(11):63-69
[7] 王力,邵明安.陕北黄土高原人工刺槐林生长与土壤干化的关系研究[J].林业科学,2004,40(1):84-91
[8] 李军,王学春,邵明安,等.黄土高原半干旱和半湿润地区刺槐林地生物量与土壤干燥化效应的模拟[J].植物生态学报,2010,34(3):330-339
[9] 方精云,陈安平.中国森林植被碳库的动态变化及其意义[J].植物学报,2001,43(9):967-973
[10]王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究[J].应用生态学报,2001,12(1) :13-16
[11]王兵,魏文俊.江西省森林碳储量与碳密度研究[J].江西科学,2007,25(6) :681-687
[12]黄从德,张健,杨万勤,等.四川人工生林态系统碳储量特征[J].应用生态学报,2008,19(8):1644-1650
[13]马钦彦,陈遐林,王娟,等.华北主要森林类型建群种的含碳率分析[J].北京林业大学学报,2002,24(5):96-100
[14]王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究[J].应用生态学报,2001,12(1):13-16
[15]刘国华,傅伯杰,方精云.中国森林碳动态及其对全球碳平衡的贡献[J].生态学报,2000,20(5):733-740
