夏蜡梅和美国蜡梅及属间杂种‘红运’营养器官解剖结构特征比较
2011-12-31史云云姚青菊徐增莱
汪 琼,史云云,姚青菊,徐增莱,吕 晔,杭 珍
〔江苏省·中国科学院植物研究所(南京中山植物园),江苏 南京 210014〕
夏蜡梅〔Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang〕隶属于蜡梅科(Calycanthaceae)夏蜡梅属(Sinocalycanthus Cheng et S.Y.Chang),特产于中国亚热带地区,自然分布于安徽省绩溪县、浙江省临安市和天台县极狭小的范围内,为中国特有的第三纪孑遗植物,属国家二级珍稀濒危保护植物。夏蜡梅一般在初夏开花,花朵洁白、硕大,花蕊金黄色,观赏价值较高[1-2]。同科的美国蜡梅属(Calycanthus L.)特产于美国东南部及加利福尼亚州,共2种2变种,美国蜡梅(C.floridus L.)为其中之一。夏蜡梅属与美国蜡梅属呈东亚—北美间断分布,在植物区系研究中具有极高的科研价值,是大陆板块漂移说强有力的佐证之一[3]。
据报道,Lasseigne等在1991年成功获得夏蜡梅和美国蜡梅的属间杂种1株,1996年该杂种植株开花[4],但未见后续的研究报道。江苏省·中国科学院植物研究所自1997年开始进行夏蜡梅和美国蜡梅的属间杂交研究,并于2001年以美国蜡梅为父本、夏蜡梅为母本,成功获得1株杂种植株;该杂种植株于2006年春初次开花,但开花数量较少;2007年春该杂种植株的开花数量增多,花大色艳,外花被红色、内花被金黄色,艳丽动人[5]。迄今为止,对该杂种已经陆续进行了优势表现、形态与光合生理特征等方面的研究[6-7],并申报品种,命名为夏蜡梅‘红运’(S.chinensis×C.floridus‘Hong Yun’)。
然而,对夏蜡梅杂种‘红运’与其亲本的解剖结构的比较研究尚无报道。为此,作者采用石蜡切片法对‘红运’及其亲本夏蜡梅和美国蜡梅营养器官横切面的解剖结构进行了观察和比较,以期为蜡梅科植物的品种选育和开发利用提供理论依据。
1 材料和方法
1.1 材料
供试的夏蜡梅杂种‘红运’及其亲本夏蜡梅和美国蜡梅均栽植于江苏省·中国科学院植物研究所实验地;于2009年5月选取生长较一致的成熟叶片、茎段和根段进行实验。
1.2 方法
用FAA溶液固定供试材料,采用常规石蜡切片法制片;用HM315切片机(德国MICROM公司生产)切片,切片厚度10~12μm,番红-固绿染色;将切片置于Nikon Eclipse 50i光学显微镜下观察和拍照。
采用Image-Pro Plus 5.0软件进行测量,每个样品测量5张片,每张片测量10个数据,结果取平均值。采用SPSS 17.0软件进行数据统计和分析。
2 结果和分析
2.1 叶横切面解剖结构特征的比较
2.1.1 共同特征 夏蜡梅、美国蜡梅和杂种‘红运’叶的横切面解剖结构有许多共同特征:均为典型的异面叶,叶片的解剖结构均可分为表皮、叶肉和叶脉3部分(图1)。
上表皮为1列圆形、椭圆形或长圆形的表皮细胞,外被薄的角质层;下表皮为1列长圆形或短圆形细胞,无角质层。叶肉分化出栅栏层和海绵层。主脉在远轴面突出,维管束为外韧型;维管束外方的中柱鞘呈环状;木质部轮廓近半圆形,其中的大型导管排列成行;韧皮部半环状,包围在木质部外方;维管束内散生大量草酸钙簇晶,外侧也存在少量针晶。
2.1.2 不同特征 叶横切面的观察结果表明:美国蜡梅和杂种‘红运’叶片的上表皮均分布有稀疏表皮毛,而夏蜡梅的叶上表皮未观察到表皮毛的存在,但其叶下表皮散生有较多的单细胞毛。比较而言,美国蜡梅的叶上表皮细胞最大、夏蜡梅叶上表皮细胞最小,杂种‘红运’叶上表皮细胞的大小与夏蜡梅接近,且二者的叶上表皮细胞大小与美国蜡梅差异显著。
杂种‘红运’和美国蜡梅叶肉的栅栏组织和海绵组织分化明显。美国蜡梅叶肉中的栅栏组织约占横切面的2/5,由2~3层长圆形或椭圆形细胞组成,表皮内侧的1层细胞排列较为紧密,其下方细胞的排列略为疏松且细胞相对较小(图1);海绵组织由5~7层圆形至椭圆形细胞组成,细胞空隙大。杂种‘红运’叶肉的栅栏组织约占横切面的1/3,细胞呈长圆形,共3~4层,排列较为紧密;海绵组织由5~7层细胞组成,细胞呈圆形至椭圆形,排列疏松,且具有较大的空腔。夏蜡梅叶肉的栅栏组织和海绵组织分化相对不十分明显(图1);栅栏组织约占横切面的1/5,由1~2层不规则细胞组成;海绵组织细胞呈圆形至短柱形,细胞层数4~5层,排列紧密,仅在气孔处形成空腔;叶肉细胞内散生有草酸钙簇晶。
杂种‘红运’及夏蜡梅和美国蜡梅的叶片横切面显微形态特征参数见表1。由表1可以看出:杂种‘红运’的叶片厚度和海绵组织厚度稍大于夏蜡梅,差异不明显,但二者的叶片厚度和海绵组织厚度均显著大于美国蜡梅。夏蜡梅的栅栏组织厚度显著小于美国蜡梅和杂种‘红运’。杂种‘红运’的主脉厚度明显大于夏蜡梅和美国蜡梅,且后两者主脉厚度相同。杂种‘红运’的叶主脉维管束横截面面积、栅栏组织厚度与海绵组织厚度的比值(P/S)和栅栏组织厚度与叶片厚度的比值(P/L)均介于其双亲夏蜡梅和美国蜡梅之间,且三者的P/S和P/L值均存在显著差异,其中,美国蜡梅的数值最高、夏蜡梅最低。三者的叶片上、下表皮厚度均以杂种‘红运’最小、美国蜡梅最大,但三者的叶上表皮厚度较为接近且无显著差异。
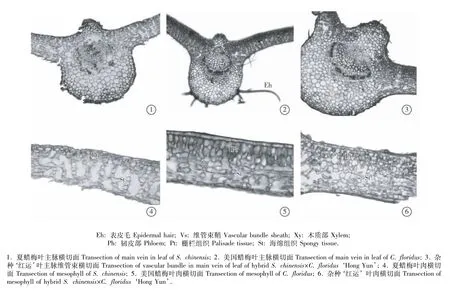
图1 夏蜡梅和美国蜡梅及属间杂种‘红运’叶片横切面解剖结构Fig.1 Transection anatom ical structure of leaf of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’
表1 夏蜡梅和美国蜡梅及属间杂种‘红运’叶片横切面解剖结构特征的比较(±SD)1)Table 1 Com parison of transection anatom ical structure characteristicsof leaf of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’(±SD)1)
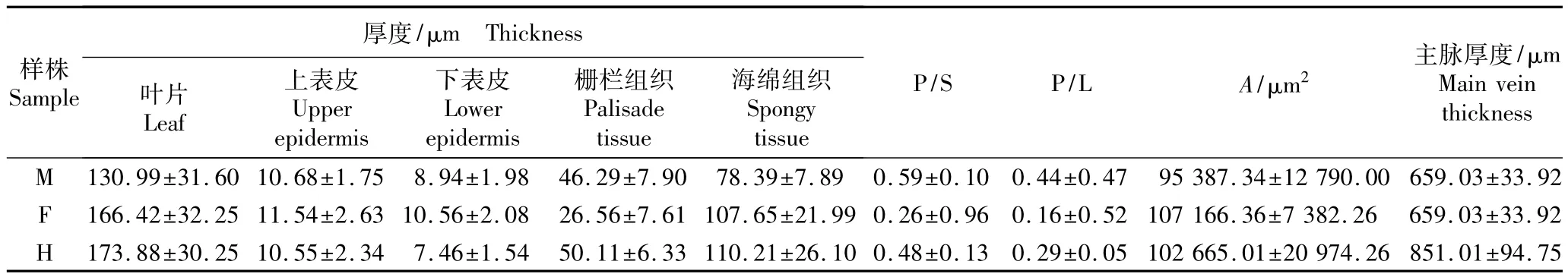
表1 夏蜡梅和美国蜡梅及属间杂种‘红运’叶片横切面解剖结构特征的比较(±SD)1)Table 1 Com parison of transection anatom ical structure characteristicsof leaf of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’(±SD)1)
1)M:父本美国蜡梅Male parent Calycanthus floridus L.;F:母本夏蜡梅Female parent Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang;H:属间杂种‘红运’Intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’;P/S:栅栏组织厚度与海绵组织厚度的比值Thickness ratio of palisade tissue to spongy tissue;P/L:栅栏组织厚度/叶片厚度的比值Thickness ratio of palisade tissue to leaf;A:主脉维管束横截面面积Transection area of vascular bundle in main vein.
样株Sample厚度/μm Thickness叶片Leaf上表皮Upper epidermis下表皮Lower epidermis栅栏组织Palisade tissue海绵组织Spongy tissue P/S P/L A/μm2 主脉厚度/μm Main vein thickness M 130.99±31.60 10.68±1.75 8.94±1.98 46.29±7.90 78.39±7.89 0.59±0.10 0.44±0.47 95 387.34±12 790.00 659.03±33.92 F 166.42±32.25 11.54±2.63 10.56±2.08 26.56±7.61 107.65±21.99 0.26±0.96 0.16±0.52 107 166.36±7 382.26 659.03±33.92 H 173.88±30.25 10.55±2.34 7.46±1.54 50.11±6.33 110.21±26.10 0.48±0.13 0.29±0.05 102 665.01±20 974.26 851.01±94.75
2.2 茎横切面解剖结构特征的比较
2.2.1 共同特征 根据夏蜡梅、美国蜡梅及属间杂种‘红运’茎横切面解剖结构的观察结果可见:三者幼茎的横切面均略呈四棱形,由外向内依次为表皮、皮层、维管柱、髓(图2)。
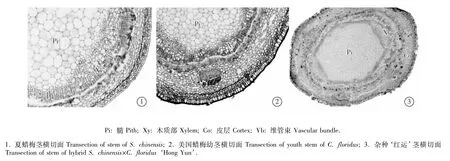
图2 夏蜡梅和美国蜡梅及属间杂种‘红运’茎横切面解剖结构Fig.2 Transection anatom ical structure of stem of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’
表皮细胞由1层圆形或扁圆形细胞组成,细胞排列紧密、整齐,细胞外壁被蜡质层和角质层覆盖。皮层中靠近表皮的数层细胞为厚角细胞,细胞较小且排列紧密;皮层内为薄壁细胞,细胞排列疏松,细胞间隙大;皮层内散生4个皮层维管束,均为木质部靠外、韧皮部在内;皮层内侧有散生的小型纤维群,排列成断续环状;纤维群的内方有小型筛管组织,形成层不明显;形成层的内方为木质部,由导管和木纤维群组成,导管细胞紧密排列成一圆环。髓位于茎的中央,由大型薄壁细胞构成,靠近外部的细胞较小,由中部向内细胞逐渐增大。
2.2.2 不同特征 观察结果表明:夏蜡梅和属间杂种‘红运’成熟茎的最外侧为木栓层,细胞不规则形,2~5层,大小不等;美国蜡梅的木栓层细胞1~2层。
美国蜡梅幼茎的木栓层内下皮层明显,由1列长椭圆状细胞组成,细胞呈垂直、紧密的径向排列;成熟茎的下皮层细胞缩短,木栓层由多层细胞构成。夏蜡梅和杂种‘红运’的茎均无明显的下皮层。夏蜡梅和杂种‘红运’的纤维群细胞成群,细胞散生并排列成断续环状,细胞极度加厚;夏蜡梅的韧皮纤维群细胞不明显,木质部较狭窄。
属间杂种‘红运’及其双亲夏蜡梅和美国蜡梅茎横切面的显微形态特征参数见表2。由表2可以看出:在1年生茎中,三者茎皮层厚度与茎半径的比值(C/R)差异不大,杂种‘红运’茎的C/R值稍高;在茎直径、皮层厚度、木质部厚度和维管束面积等方面三者差异明显,其中,杂种‘红运’茎的这几项指标的数值均最高。在当年生幼茎中,夏蜡梅和美国蜡梅茎横切面中髓面积所占比例较接近,明显高于杂种‘红运’;而在1年生茎中,三者茎横切面中髓面积所占比例差异不大,其中,夏蜡梅稍高于美国蜡梅和杂种‘红运’。
2.3 根横切面解剖结构特征的比较
2.3.1 共同特征 夏蜡梅、美国蜡梅及属间杂种‘红运’根横切面的解剖结构具有很多相同特征,三者根横切面均为近圆形,由外向内依次为木栓层、皮层、石细胞环、韧皮部和木质部(图3)。
表2 夏蜡梅和美国蜡梅及属间杂种‘红运’茎横切面解剖结构特征的比较(±SD)1)Table 2 Com parison of transection anatom ical structure characteristics of stem of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’(±SD)1)

表2 夏蜡梅和美国蜡梅及属间杂种‘红运’茎横切面解剖结构特征的比较(±SD)1)Table 2 Com parison of transection anatom ical structure characteristics of stem of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’(±SD)1)
1)M:父本美国蜡梅Male parent Calycanthus floridus L.;F:母本夏蜡梅Female parent Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang;H:属间杂种‘红运’Intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’;C/R:皮层厚度与茎半径的比值 Ratio of cortex thickness to stem radius;P1:髓面积占1年生茎横截面总面积的比例Proportion of pith area in total area of transection of one-year-old stem;P2:髓面积占当年生幼茎横截面总面积的比例Proportion of pith area in total area of transection of current year youth stem.
样株Sample茎直径/μm Stem diameter皮层细胞层数Layer number of cortex cell皮层厚度/μm Cortex thickness C/R 木质部厚度/μm Xylem thickness维管束面积/μm2 Vascular bundle area P1 P2 M 2 011.92±259.94 10-13 262.67±21.03 0.25±0.03 220.12±57.96 42 090.24±95 831.96 0.24±0.01 0.48±0.78 F 2 342.24±423.71 8-12 277.70±59.55 0.24±0.02 211.44±34.39 44 087.07±16 281.10 0.28±0.02 0.45±0.62 H 2 852.43±408.64 11-13 374.12±41.80 0.26±0.02 305.80±29.33 51 422.53±19 702.27 0.21±0.05 0.29±0.04
木栓层多由2层栓化加厚的长圆形或近圆形细胞组成,最外层部分脱落。皮层由7~9层薄壁细胞组成,其中,外侧的2~3层细胞呈长圆形,排列紧密,细胞间隙小;内侧有5~6层细胞,形状不规则,排列疏松,细胞间隙大。在皮层内侧、韧皮部外方有1~2层石细胞环。韧皮部由薄壁细胞和筛管细胞组成。木质部包含木射线和导管。皮层和韧皮部细胞中一般有草酸钙晶体和淀粉粒等后含物。
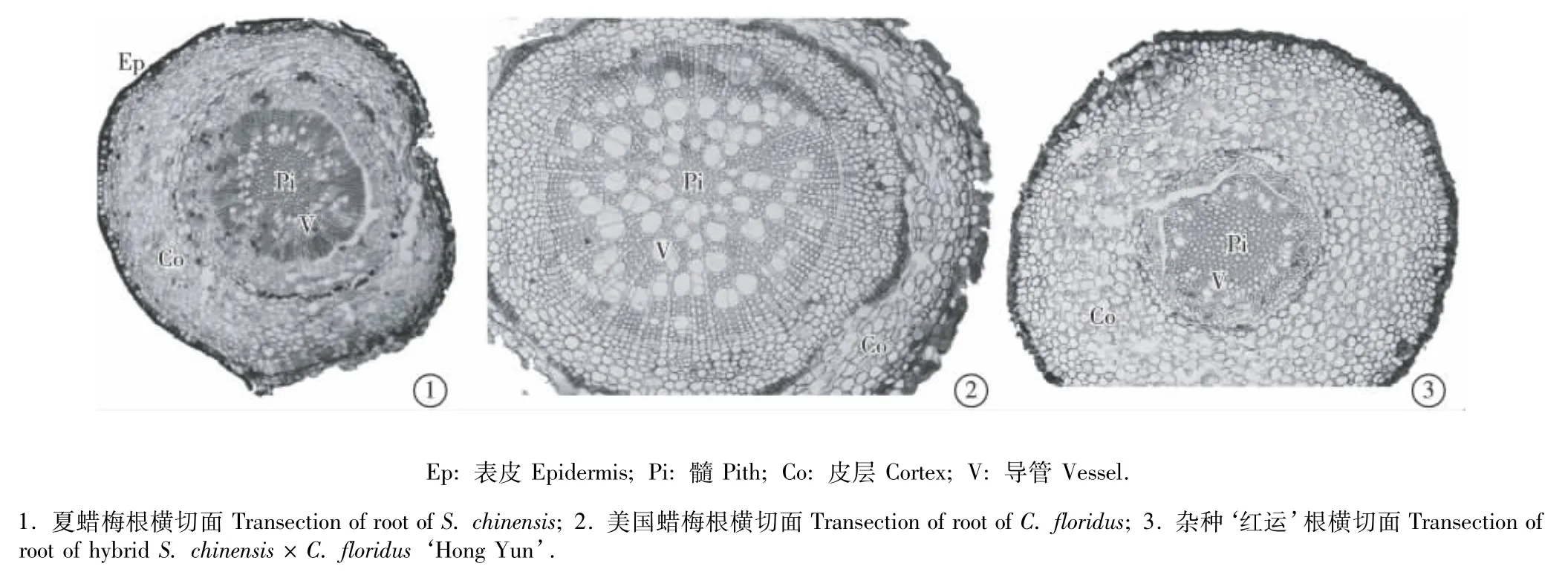
图3 夏蜡梅和美国蜡梅及属间杂种‘红运’根横切面解剖结构Fig.3 Transection anatom ical structure of root of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’
2.3.2 不同特征 观察结果表明:三者根的解剖结构特征也有一定的差异。美国蜡梅和杂种‘红运’根木栓层的最外侧细胞小而圆,而夏蜡梅的则呈长圆形、切向排列。夏蜡梅根皮层的最外侧有1~2层石细胞环,最内侧也有同样的石细胞环(图3-1);美国蜡梅根皮层最外侧的1~3层细胞部分石细胞化并形成断续环,最内侧也有类似的石细胞环(图3-2);杂种‘红运’根皮层的最外侧无石细胞环,只在最内侧有石细胞构成的断续环(图3-3)。美国蜡梅根皮层最窄;杂种‘红运’根的韧皮部较夏蜡梅和美国蜡梅狭窄。夏蜡梅根木质部的木射线呈放射状排列,美国蜡梅根木质部的薄壁细胞分散着较多的大型导管。杂种‘红运’根木质部轮廓呈五边形,而其双亲根木质部的轮廓则为近圆形。夏蜡梅和杂种‘红运’根的中央均有明显的髓,由木栓化加厚的细胞组成;美国蜡梅根的髓不明显。美国蜡梅根木质部含有较多直径较大的导管,杂种‘红运’根木质部所含导管数量最少。夏蜡梅根皮层中的细胞所含淀粉粒较少,但韧皮部细胞含较多淀粉粒;美国蜡梅根皮层和韧皮部细胞中均含较多淀粉粒;杂种‘红运’根皮层细胞含丰富的淀粉粒,但韧皮部细胞中淀粉粒却较少。夏蜡梅根韧皮部中有大量的簇晶,其皮层中也有较多的簇晶;美国蜡梅和杂种‘红运’根韧皮部中均有簇晶分布,但二者根皮层中却少见簇晶。比较而言,美国蜡梅根髓部大型导管最多、杂种‘红运’最少。
杂种‘红运’及其双亲夏蜡梅和美国蜡梅根横切面的显微形态特征参数见表3。三者根木栓层厚度较为接近,差异不显著。夏蜡梅根直径最大、杂种‘红运’最小。三者髓面积在根横截面总面积中所占比例差异很大;其中美国蜡梅最小,平均比例为夏蜡梅的14%、为杂种‘红运’的7%。三者根的皮层厚度、皮层厚度与根半径的比值(C/R)和髓面积占根横截面总面积的比例这3个参数差异显著,且均以杂种‘红运’的数值最高、美国蜡梅最小。三者根的木质部厚度差异很大,美国蜡梅根木质部的厚度为杂种‘红运’的2.95倍、夏蜡梅的1.74倍。根木质部厚度与根半径的比值(X/R)也是美国蜡梅最高,夏蜡梅和杂种‘红运’根的X/R值则较为接近。
表3 夏蜡梅和美国蜡梅及属间杂种‘红运’根横切面解剖结构特征的比较(±SD)1)Table 3 Com parison of transection anatom ical structure characteristics of root of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’(±SD)1)

表3 夏蜡梅和美国蜡梅及属间杂种‘红运’根横切面解剖结构特征的比较(±SD)1)Table 3 Com parison of transection anatom ical structure characteristics of root of Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang,Calycanthus floridus L.and intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’(±SD)1)
1)M:父本美国蜡梅Male parent Calycanthus floridus L.;F:母本夏蜡梅Female parent Sinocalycanthus chinensis(Cheng et S.Y.Chang)Cheng et S.Y.Chang;H:属间杂种‘红运’Intergeneric hybrid S.chinensis×C.floridus‘Hong Yun’;C/R:皮层厚度与根半径的比值 Ratio of cortex thickness to root radius;X/R:木质部厚度与根半径的比值 Ratio of xylem thickness to root radius;P1:髓面积占根横截面总面积的比例Proportion of pith area in total area of transection of root.
样株Sample厚度/μm Thickness木栓层Phellem 皮层Cortex 木质部Xylem皮层细胞层数Layer number of cortex cell根直径/μm Root diameter C/R X/R P1 大型导管数Number of big vessel M 28.20±3.09 161.69±29.77 326.85±58.46 7-9 1 437.73±179.18 0.20±0.67 0.46±0.07 0.005±0.00 30-63 F 23.39±2.81 312.90±52.28 187.51±56.10 10-11 1 718.99±185.18 0.37±0.09 0.22±0.04 0.035±0.01 20-50 H 26.75±4.07 316.20±24.82 110.72±14.69 10-14 1 074.17±67.45 0.59±0.05 0.21±0.02 0.071±0.10 14-20
3 讨论和结论
夏蜡梅、美国蜡梅和杂种‘红运’在叶片横切面的显微结构上有一定的差异,主要表现在叶肉的栅栏组织和海绵组织的分化程度等方面。杂种‘红运’的叶片厚度、主脉维管束横截面面积和海绵组织厚度更接近于夏蜡梅,与美国蜡梅差异较大。杂种‘红运’叶的栅栏组织厚度与海绵组织厚度的比值(P/S)和栅栏组织厚度与叶片厚度的比值(P/L)均介于双亲之间。研究表明[8-11]:P/S和P/L值可以作为评价植株抗旱性、抗寒性和光合能力的重要指标。观测结果表明:美国蜡梅叶的P/S和P/L均最高,杂种‘红运’其次,夏蜡梅最低;夏蜡梅叶的P/L仅为0.16,在一定程度上证明夏蜡梅具有不喜强光、稍耐阴的生态特性[12]。从叶解剖结构特征上看,杂种‘红运’在耐热性和光合能力方面均优于夏蜡梅,而美国蜡梅则优于夏蜡梅和杂种‘红运’,这与盛宁等[7]的研究结果一致。
在茎解剖结构方面,杂种‘红运’的茎直径、皮层细胞层数、皮层厚度、皮层厚度与茎半径的比值、木质部厚度和维管束面积等指标均高于其双亲美国蜡梅和夏蜡梅,而杂种‘红运’茎的髓面积在其1年生和当年生幼茎横截面中所占比例却最小。
杂种‘红运’根的解剖结构与其双亲存在一定差异。美国蜡梅根的髓部不发达,而夏蜡梅根则有明显的髓部,杂种‘红运’根的髓部较夏蜡梅更发达。美国蜡梅根的木质部和木栓层均最厚,髓中的大型导管数量最多;而杂种‘红运’根的皮层最发达,木质部最不发达,髓中的大型导管数量也最少。
总体上看,夏蜡梅、美国蜡梅和属间杂种‘红运’营养器官的主要解剖结构特征均表现出较高的共性,但仍存在一定差异。其中,叶片显微结构差异最大,这可能是因为叶片长期暴露于外界多变的环境中,与周围环境发生的物质和能量交流最为频繁,因此表现出很强的变异性和可塑性。
[1]郑万钧,章绍尧.蜡梅科的新属——夏蜡梅属[J].植物分类学报,1964,9(2):135-138.
[2]傅立国,金鉴明.中国植物红皮书——稀有濒危植物:第一册[M].北京:科学出版社,1991:196-197.
[3]张若蕙,刘洪谔.世界蜡梅[M].北京:中国科学技术出版社,1998.
[4]Lasseigne F T,Fantz P R,Raulston J C,et al.×Sinocaly-calycanthus raulstonii(Calycanthaceae):a new intergeneric hybrid between Sinocalycanthus chinensis and Calycanthus floridus[J].HortScience,2001,36(4):765-767.
[5]姚青菊,夏 冰,蔡小龙,等.夏蜡梅和美国蜡梅属间杂种培育成功[J].江苏农业科学,2007(3):222.
[6]姚青菊,夏 冰,任全进,等.夏蜡梅和美国蜡梅属间杂种的优势表现[J].江苏林业科技,2007,34(4):24-26,35.
[7]盛 宁,姚青菊,任全进,等.夏蜡梅和美国蜡梅属间杂种形态与光合生理特征[J].浙江林学院学报,2008,25(6):728-732.
[8]贺金生,陈伟烈,王勋陵.高山栎叶的形态结构及其与生态环境的关系[J].植物生态学报,1994,18(3):219-227.
[9]王 玉,洪永聪,丁兆堂,等.利用茶树叶片解剖结构指数预测茶树种质材料的抗寒性[J].中国农学通报,2009,25(9): 126-130.
[10]张惠斌,刘星辉.龙眼叶片组织细胞结构特性与耐寒性的关系[J].园艺学报,1993,20(1):1-7.
[11]张红瑞,高致明,李志敏.牛膝两个农家品种光合特性及叶片解剖结构的比较[J].中国农学通报,2008,24(12):142-145.
[12]陈模舜,柯世省.濒危植物夏蜡梅营养器官的解剖结构特征[J].植物资源与环境学报,2010,19(3):37-41.
