华山新麦草叶片的光合生理特性
2011-12-31柴永福张晓飞
柴永福,李 倩,张晓飞,岳 明
(西北大学西部资源生物与现代生物技术教育部重点实验室,陕西 西安 710069)
华山新麦草(Psathyrostachys huashanica Keng ex P.C.Kuo)为多年生草本植物,隶属于禾本科(Poaceae)新麦草属(Psathyrostachys Nevski),是中国特有种,主要分布在陕西华山的华山峪、黄浦峪和仙峪[1]。华山新麦草是小麦属(Triticum L.)植物的野生近缘种,具有抗病、抗旱和早熟等特点,在探索小麦属植物的起源、进化及遗传育种等方面具有重要价值[2],已被列为国家一级珍稀濒危植物和急需保护的农作物野生亲缘种[3]。
近年来,对华山新麦草的研究主要集中在系统进化、杂交育种和生殖生物学等方面[4],而关于华山新麦草生理生态学方面的研究尚未见报道。为了有效保护华山新麦草这一野生种质资源、探讨其致濒机制,作者对华山新麦草的光合生理特性进行了分析。
1 自然分布现状和研究方法
1.1 自然分布现状
目前,华山新麦草仅在陕西省华阴市境内的华山有分布。华山是秦岭支脉华山山脉的一部分,地理坐标为北纬34°25′~34°30′、东经109°57′~110°05′;该区域属大陆性季风气候,四季分明,季节气温差别较大,冬季寒冷干燥、夏季炎热多雨;年均降雨量902.7 mm,年均气温5.9℃,最高温度27.7℃,最低温度-24.1℃。华山新麦草分布的海拔范围较广,从山麓平原(海拔351 m)到山顶(海拔2 153 m)均有分布,海拔高差1 800 m[5];该种在向阳地段分布较密集,在背光处分布较稀疏,在林下或阴湿处未见分布;喜干旱较温暖气候,在降雨较少的中低山地区及山麓地带分布数量相对较多,而在高山地带分布数量相对较少[6];多生长在岩石凹陷或空隙处;生长的土壤主要是微生物分解土层表面腐败的植物脱落或死亡器官形成的腐殖质,土壤中的水分主要依靠降水,因此,降水的不确定性导致了华山新麦草分布区域土壤含水量的不规律性[5]。
1.2 材料和方法
供试华山新麦草采自华山黄浦峪并引种栽培于西安市西北大学生物系果园内,株高约40~50 cm。栽培所用土壤均来源于华山新麦草原生长地,栽培期间每隔3 d浇水1次。
于2010年4月上旬在晴朗无云的天气条件下随机选择健康、正常的华山新麦草营养叶5片,用美国CID公司生产的CI-301PS便携式光合测定仪测定净光合速率、蒸腾速率、叶温、气孔阻力、光合有效辐射强度和气温等相关指标。测定时系统处于开放状态,测定时间为每天7:30至19:30,1 h测量1次,重复测量5 d,结果取平均值。
1.3 数据处理与分析
运用Statistic 6.0软件对实验数据进行统计分析,采用最小二乘法对光响应方程进行拟合,日变化曲线和光响应曲线均使用Origin 7.5软件完成。
2 结果和分析
2.1 净光合速率的日变化
华山新麦草净光合速率日变化曲线见图1。与大多数植物净光合速率的日变化曲线呈“双峰”型的趋势有明显不同[7-11],华山新麦草净光合速率的日变化曲线呈“三峰”型,共包含有3个峰值,3个峰值出现的时间分别为9:30、11:30和16:30,对应的净光合速率分别为6.5、6.2和9.0μmol·m-2·s-1,第2个峰和第3个峰之间出现“午降”现象,“午降”时(中午12:30左右)华山新麦草叶片的净光合速率较低,仅为2.5μmol·m-2·s-1。
2.2 蒸腾速率的日变化
蒸腾作用的强弱是衡量植物水分代谢能力的重要生理指标[12]。华山新麦草蒸腾速率的日变化曲线见图2。结果表明:华山新麦草蒸腾速率的日变化曲线呈“单峰”型,峰值为1.7 mmol·m-2·s-1,出现在13:30;在11:30至13:30,叶片蒸腾速率的上升速率最快;13:30后叶片的蒸腾速率逐渐下降,在19:30下降至0.3 mmol·m-2·s-1。
2.3 叶温日变化及其对净光合速率的影响
华山新麦草叶温的日变化曲线以及叶温与其净光合速率的关系图分别见图3和图4。华山新麦草叶温的日变化曲线呈“单峰”型(图3),在12:30达到最大值(39.8℃)。根据叶温与净光合速率的变化拟合得到华山新麦草净光合速率对叶温的响应方程y= -0.023 78x2+1.371 18x-13.123 50(r2=0.353 9)。由图4可见:在一定叶温范围内,净光合速率与叶温呈正相关,随着叶温的升高净光合速率逐渐增大,当叶温达到28℃时净光合速率最大,但叶温超过28℃时净光合速率开始逐渐下降。

图1 华山新麦草净光合速率的日变化曲线Fig.1 Daily change curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo
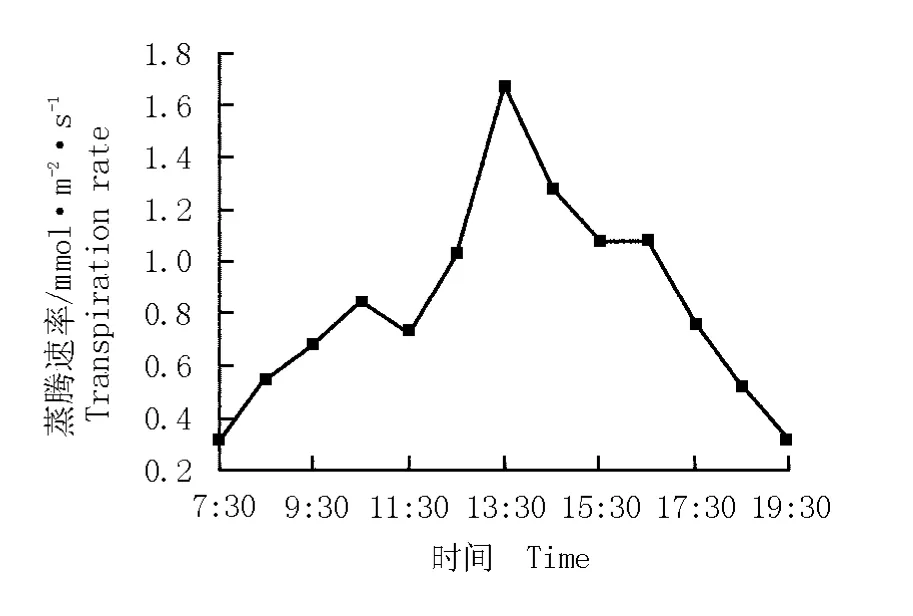
图2 华山新麦草蒸腾速率的日变化曲线Fig.2 Daily change curve of transpiration rate of Psathyrostachys huashanica Keng ex P.C.Kuo
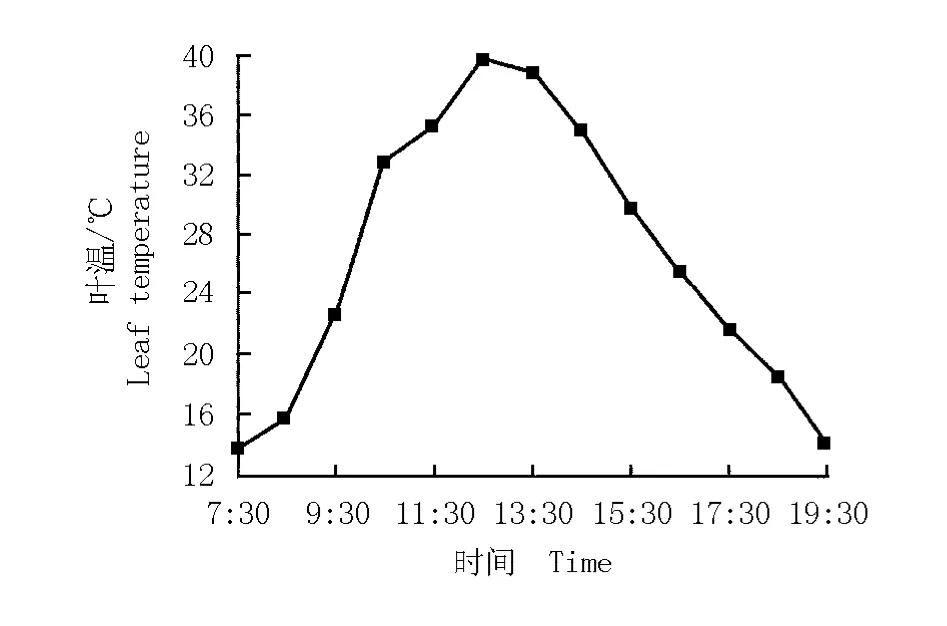
图3 华山新麦草叶温的日变化曲线Fig.3 Daily change curve of leaf tem perature of Psathyrostachys huashanica Keng ex P.C.Kuo

图4 华山新麦草净光合速率对叶温的响应曲线Fig.4 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to leaf tem perature
2.4 气孔阻力日变化及其对净光合速率的影响
华山新麦草气孔阻力的日变化曲线以及气孔阻力与其净光合速率的关系图分别见图5和图6。结果表明:华山新麦草气孔阻力的日变化曲线呈“单峰”型,在11:30达到最大值,为74 m2·s·mol-1。根据气孔阻力与净光合速率的变化拟合得到华山新麦草净光合速率对气孔阻力的响应方程y=0.000 37x2-0.104 19x+7.895 66(r2=0.510 6)。由图6可见:华山新麦草的净光合速率与气孔阻力呈负相关关系,随着气孔阻力的增大,净光合速率急速下降。
2.5 光合有效辐射强度日变化及其对净光合速率的影响
华山新麦草生境中光合有效辐射强度的日变化曲线及其与净光合速率的关系图分别见图7和图8。由图7可看出:华山新麦草生境中光合有效辐射强度的日变化曲线呈“单峰”型。从7:30开始,光合有效辐射强度迅速增大,10:30达到600μmol·m-2·s-1,12:30达到最大值(900μmol·m-2·s-1);随后,光合有效辐射强度逐渐降低,至19:30几乎接近于0μmol·m-2·s-1。
根据光合有效辐射强度与净光合速率的变化拟合得到华山新麦草净光合速率对光合有效辐射强度的响应方程y=-0.000 03x2+0.031 89x-0.035 49 (r2=0.669 0)。由图8可见:华山新麦草的净光合速率随光合有效辐射强度增大而呈“抛物线”型变化,光合有效辐射强度在300~700μmol·m-2·s-1范围内净光合速率较高;净光合速率达到最高时的光合有效辐射强度为500μmol·m-2·s-1。将叶片净光合速率为0μmol·m-2·s-1时的光照强度作为光补偿点(light compensation point,LCP),将净光合速率最高时的拐点作为光饱和点(light saturation point,LSP),根据拟合方程计算得出华山新麦草的LCP和LSP分别为1.1和531.5μmol·m-2·s-1。
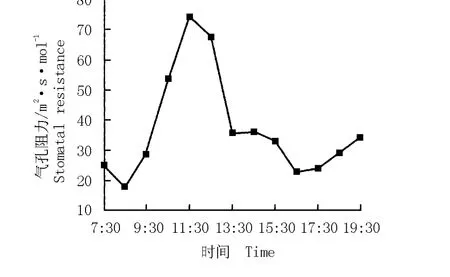
图5 华山新麦草气孔阻力的日变化曲线Fig.5 Daily change curve of stomatal resistance of Psathyrostachys huashanica Keng ex P.C.Kuo

图6 华山新麦草净光合速率对气孔阻力的响应曲线Fig.6 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to stomatal resistance

图7 华山新麦草生境中光合有效辐射强度(PAR)的日变化曲线Fig.7 Daily change curve of photosynthetically active radiation intensity(PAR)in habitat of Psathyrostachys huashanica Keng ex P.C.Kuo

图8 华山新麦草净光合速率对光合有效辐射强度(PAR)的响应曲线Fig.8 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to photosynthetically active radiation intensity(PAR)
2.6 气温日变化及其对净光合速率的影响
气温与光合有效辐射强度常常是密切相关的,光合有效辐射强度大,气温也相应较高,因而,气温与植物的净光合速率也具有相关性。华山新麦草生境中气温的日变化曲线及其与净光合速率的关系图分别见图9和图10。图9显示:华山新麦草生长环境的气温日变化曲线也呈“单峰”型,从7:30开始气温持续升高;10:30气温已达到26.8℃;12:30至14:30间的气温较高,保持在32℃以上;13:30气温达到峰值,随后逐渐下降。
根据华山新麦草净光合速率与气温的变化拟合得到华山新麦草净光合速率对气温的响应方程y= -0.138 89x2+7.162 14x-85.225 39(r2=0.394 0)。由图10可见:华山新麦草净光合速率对气温的响应曲线也呈“抛物线”型,与其对光合有效辐射强度的响应曲线相似。在一定范围内(低于26℃),随着气温的升高,净光合速率增大;气温超过26℃,净光合速率逐渐下降。对于华山新麦草而言,适宜的气温为20℃~30℃,最适气温为26℃,在此气温条件下华山新麦草的净光合速率可达到最高。

图9 华山新麦草生境中气温的日变化曲线Fig.9 Daily change curve of air tem perature in habitat of Psathyrostachys huashanica Keng ex P.C.Kuo
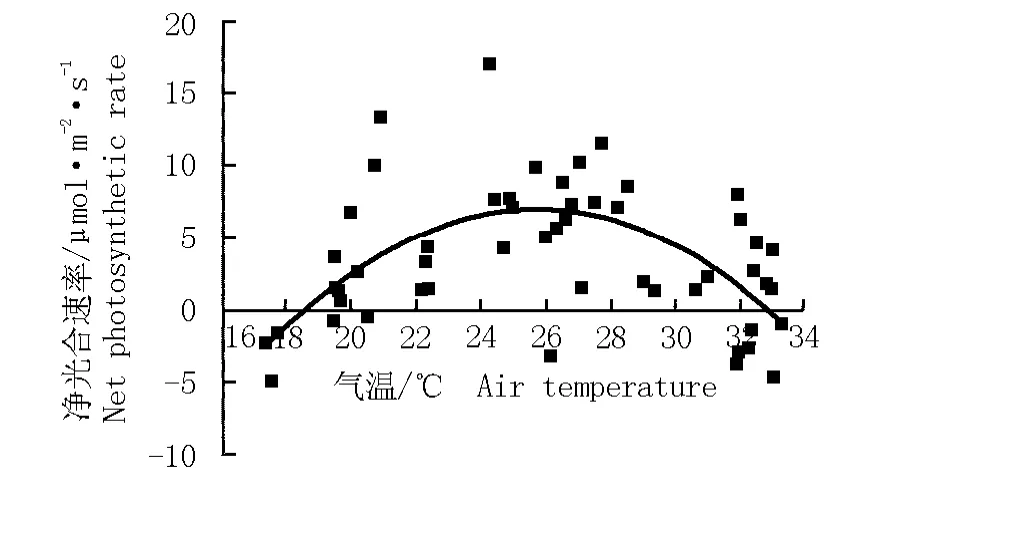
图10 华山新麦草净光合速率对气温的响应曲线Fig.10 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to air temperature
3 讨论和结论
研究结果表明:华山新麦草净光合速率的日变化曲线共存在3个峰值且数值均较小,分别为6.5、6.2和9.0μmol·m-2·s-1。灰绿型羊草〔Aneurolepidium chinense(Trin.)Kitag.〕的净光合速率日变化曲线有2个峰值,分别为23.2和17.3μmol·m-2·s-1[13];小麦(Triticum aestivum L.)的最大净光合速率为27.7 μmol·m-2·s-1[14];水稻(Oryza sativa L.)的最大净光合速率为23.6μmol·m-2·s-1[15],均高于华山新麦草的净光合速率。产生这一现象的原因有二:一是实验过程中气候干旱,华山新麦草植株处于水分胁迫状态,因而净光合速率的测定值均偏低;二是与华山新麦草本身的生理代谢特性有关。岳明等[5]的研究结果表明:华山新麦草的生存竞争能力较弱。本研究结果从净光合速率角度也印证了这个结论。
华山新麦草的净光合速率具有“午降”现象,“午降”时净光合速率的最低值为2.5μmol·m-2·s-1,明显低于灰绿型羊草“午降”时净光合速率的最低值(6.0μmol·m-2·s-1)[13]。推测华山新麦草的净光合速率在12:30左右产生“午降”现象,可能的原因为:此时的光合有效辐射强度达到全天的最大值,空气和叶片温度均较高,致使气孔关闭,进而抑制植株的光合作用;此时植株体内的光合产物已积累到一定程度,更重要的是叶内水分含量在中午最少,使植株的光合作用受到水分胁迫的影响;另外,CO2自根向地上部分的运输量在中午增加、过度光照引起光氧化反应以及呼吸作用的日变化所引发的光合作用的变动[16]等都可能与华山新麦草净光合速率的“午降”现象有关。
研究结果表明:华山新麦草叶片的蒸腾速率一直在1.7 mmol·m-2·s-1以下,明显低于灰绿型羊草蒸腾速率的“双峰”值(6.1和7.2 mmol·m-2·s-1)[13],这可能与华山新麦草叶片气孔阻力较大有关。
根据华山新麦草净光合速率对光合有效辐射强度的拟合方程,计算得出华山新麦草的光补偿点和光饱和点分别为1.1和531.5μmol·m-2·s-1。在光饱和点以下,光合有效辐射强度是华山新麦草光合作用的主要限制因素,其净光合速率随着光合有效辐射强度的增强而提高;但光合有效辐射强度达到光饱和点后,光合有效辐射强度就不再是华山新麦草光合作用的主要限制因素。植物叶片的光饱和点与光补偿点反映了植物对光照条件的要求,分别体现了对强光和弱光的利用能力。光补偿点较低、光饱和点较高的植物对光照强度的适应范围较广;而光补偿点较高、光饱和点较低的植物则对光照强度的适应范围较窄[17]。华山新麦草的光补偿点较低而光饱和点较高,显示其具有很强的喜光性,且对光照强度的适应范围较广。
气孔阻力能够反映气孔开启变化的情况,直接影响植物的光合作用和蒸腾作用[18]。华山新麦草的气孔阻力变化非常大,从早上8:30的15 m2·s·mol-1迅速增大到11:30的74 m2·s·mol-1,严重阻碍了光合作用的顺利进行。禾本科植物的气孔与其他植物不同,不但在白天仍处于半开状态,而且对光的反应很敏感。由此可以认为:在阴天和雨天或是早晨、晚上,禾本科植物的光合作用不仅受到光照条件的限制,同时还受到气孔开启程度的制约[19]。因此,气孔阻力是对华山新麦草净光合速率和蒸腾速率具有负效应的重要因子。
综合分析结果表明:华山新麦草的净光合速率与气孔阻力、光合有效辐射强度、叶温和气温等因素均有很大的关系,其中,叶温、光合有效辐射强度和气温对净光合速率的影响曲线基本为“抛物线”型,气孔阻力对净光合速率的影响则表现为“随气孔阻力的增大净光合速率持续降低”的趋势。较大的气孔阻力阻碍了华山新麦草的光合作用,使其不能有效合成代谢所需的大量营养物质,以维持自身的正常生命活动,这可能是华山新麦草濒临灭绝的主要生理原因之一。
[1]中国科学院西北植物研究所.秦岭植物志:第1卷第1分册[M].北京:科学出版社,1976:99.
[2]孙根楼,颜 济,杨俊良.普通小麦和新麦草属间杂种的产生及细胞遗传学研究[J].遗传学报,1992,19(4):322-326.
[3]卢宝荣.小麦族遗传资源的多样性及其保护[J].生物多样性,1995,3(2):63-68.
[4]王 丽,赵桂仿.华山新麦草胚和胚乳的发育研究[J].西北植物学报,2002,22(4):786-790,T005-T006.
[5]岳 明,张林静,马 凯,等.华山新麦草濒危原因及种群繁殖对策[J].生态学报,2001,21(8):1314-1320.
[6]赵利锋,李 珊,潘 莹,等.华山新麦草自然居群沿海拔梯度的遗传分化[J].西北植物学报,2001,21(3):391-400.
[7]陈德祥,李意德,骆土寿,等.海南岛尖峰岭热带山地雨林下层乔木中华厚壳桂光合生理生态特性的研究[J].林业科学研究,2003,16(5):540-547.
[8]刘志龙,方建民,虞木奎,等.三种林-茶复合林分中环境因子和茶的光合特征参数的日变化规律[J].植物资源与环境学报,2009,18(2):62-67.
[9]谭会娟,周海燕,李新荣,等.珍稀濒危植物半日花光合作用日动态变化的初步研究[J].中国沙漠,2005,25(2):262-267.
[10]王建丽,张永亮,朱占林,等.杂花苜蓿叶片光合生理生态特性[J].草地学报,2006,14(2):138-141.
[11]贾宏涛,赵成义,盛 钰,等.干旱地区紫花苜蓿光合日变化规律研究[J].草业科学,2009,26(7):56-60.
[12]邹学校,马艳青,张竹青,等.环境因子对辣椒光合与蒸腾特性的影响[J].植物资源与环境学报,2005,14(4):15-20.
[13]王德利,王正文,张喜军.羊草两个趋异类型的光合生理生态特性比较的初步研究[J].生态学报,1999,19(6):837-843.
[14]王焕忠,李雁鸣,张建平,等.不同熟期小麦品种光合性能的初步研究[J].河北农业大学学报,1998,21(2):1-5.
[15]曹树青,翟虎渠,张红生,等.不同类型水稻品种叶源量及有关光合生理指标的研究[J].中国水稻科学,1999,13(2): 91-94.
[16]徐 江,朱建雯,张巨松,等.田间棉花叶片呼吸作用对光合日变化的影响[J].新疆农业大学学报,1997,20(4):1-4.
[17]武维华.植物生理学[M].1版.北京:科学出版社,2003: 168-170.
[18]刘贞琦,伍贤进,刘振业.土壤水分对烟草光合生理特性影响的研究[J].中国烟草学报,1995,2(3):44-49.
[19]户刈义次,山田登,杉山直仪,等.作物的生理生态[M].崔继林,刘中宣,郑丕尧,等,译.北京:科学出版社,1962:27-56.
