受褐飞虱诱导的水稻酵母双杂交文库的构建
2011-11-18祝莉莉胡亮杜波
祝莉莉 胡亮 杜波

摘要:为了研究水稻抗褐飞虱的分子机制,构建了受褐飞虱取食诱导的水稻的酵母双杂交文库。检测结果表明构建的文库容量为2.22×106 CFU,文库滴度为5.4×107 CFU/mL,文库cDNA插入片段长度主要分布在700~1 000 bp之间。
关键词:褐飞虱;水稻;酵母双杂交文库
中图分类号:Q812文献标识码:A文章编号:0439-8114(2011)15-3201-03
Construction of Yeast Two-hybrid Library of Rice Infested by Brown Planthopper
ZHU Li-li,HU Liang,DU Bo
(School of Life Science, Wuhan University, Wuhan 430072, China)
Abstract: To investigate the molecular resistance mechanism of rice to brown planthopper, yeast two-hybrid library of rice infested by the insect was constructed. The results of detection showed that the library contained 2.22×106 independent clones; the titer of library were 5.4×107 CFU/mL. The size of most inserts were 700~1 000 bp in the library.
Key words: brown planthopper; rice; yeast two-hybrid library
褐飞虱(Nilaparvata lugens St?覽l.,BPH)是对水稻危害最严重的害虫之一,严重威胁水稻的高产和稳产[1]。在水稻生产中,依靠化学杀虫剂防治褐飞虱不仅可能引起褐飞虱的再猖獗,还会造成环境污染[2]。目前认为,培育并推广种植抗褐飞虱水稻品种是综合防治褐飞虱最为经济有效和环境友好的方法[3]。迄今为止,人们在野生稻和栽培品种中定位了21个抗褐飞虱的主效基因[4],并成功地克隆了水稻抗褐飞虱基因Bph14[5],但水稻抗褐飞虱的分子机制仍不清楚。近年来,科学家们应用多种方法,如cDNA array、抑制差减杂交、cDNA Microarry以及iTRAQ技术等,从DNA、RNA和蛋白质水平对抗、感水稻品种受褐飞虱取食诱导后的变化进行了探索[6-9]。这些研究表明,褐飞虱的取食导致了水稻体内与细胞防御、细胞传递、大分子代谢、信号传导等相关的基因和蛋白表达显著的改变。然而,这些报道都是大范围地进行水稻抗褐飞虱分子机制的探索。抗褐飞虱相关基因与哪种蛋白相互作用,从而介导抗性反应则完全不清楚。
酵母双杂交技术是一种研究蛋白质之间相互作用的有效手段,不但可以检测已知蛋白质之间的相互作用,更重要的在于发现与已知蛋白相互作用的未知蛋白,是一种成熟的研究蛋白间相互作用的方法。本研究构建褐飞虱取食水稻品种9311的酵母双杂交文库,为筛选抗褐飞虱基因互作蛋白分子,并进一步研究水稻抗褐飞虱的分子机理奠定基础。
1材料和方法
1.1实验材料
建库所用水稻品种为9311;所用褐飞虱饲养在武汉大学植物遗传研究所的水稻品种台中本地一号(TN1)上。构建酵母文库的试剂盒购自Clontech公司,RNA提取试剂盒购自Invitrogene公司。
1.2方法
1.2.1水稻处理将水稻品种9311播种于直径9 cm的塑料杯中。4叶期时分别于0、3、6、12、24、48、72、96 h按每棵苗8头间隔接入褐飞虱,同一时间终止反应。
1.2.2RNA的提取采用Invitrogen公司的RNA提取试剂盒提取水稻叶鞘的RNA。提取后的RNA进行电泳,检测其质量,并调整RNA的量至所有样品相同。
1.2.3cDNA的合成参照Clontech试剂盒的说明,先合成cDNA第一链,再通过PCR扩增双链cDNA,并纯化以去除小片段cDNA。
1.2.4酵母感受态细胞的制备按照Clontech试剂盒的说明制备Y187感受态细胞,制备好的感受态细胞要立即使用。
1.2.5酵母文库的转化按照Clontech试剂盒的要求,将扩增的双链cDNA、线性化的pGADT7-Rec转入酵母感受态细胞Y187。在直径150 mm的SD/-Leu培养皿上涂抹150 μL菌液(共100个培养皿)。在直径90 mm的SD/-Leu培养皿上分别涂抹1∶10,1∶100和1∶1 000的稀释菌液,以检测转化的效率。培养皿在30 ℃条件下倒置培养3 d至菌落出现。
1.2.6文库的收集和滴度测定根据按比例稀释的平板上的克隆数,计算文库的独立转化的克隆:初始文库独立克隆子数目(库容量)=每块平板的平均克隆子数目×稀释倍数×悬浮体积。然后用含有30%甘油的LB液体培养基清洗培养基上生长的菌落,全部收集于无菌三角瓶中,共300 mL。充分混匀后,用血球计数板进行滴度测定。符合滴度要求,再按每1 mL分装至1.5 mL的EP管,保存在-80 ℃。
1.2.7cDNA插入片段大小的检测随机挑取平板上生长的单菌落,以pGADT7载体通用引物T7和AD对其进行PCR扩增,电泳检测cDNA插入片段的大小。
2结果与分析
2.1RNA的质量及浓度
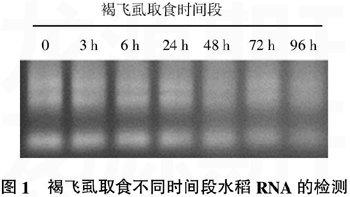
用紫外分光光度计和琼脂糖凝胶电泳检测提取的总RNA纯度(图1)。结果显示:提取的RNA含量高,纯度好,未发生降解,满足建库需要;同时,调整RNA样品的上样量,使受褐飞虱取食8个时间段的水稻RNA样品量相同。相同质量的RNA混合后,反转录得到单链cDNA。
2.2cDNA合成和纯化结果
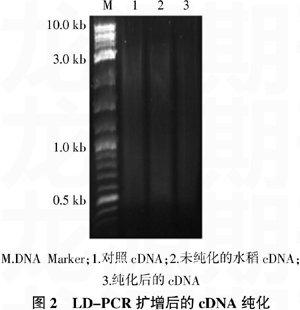
mRNA反转录合成第一链后,LD-PCR扩增获得双链cDNA。取5 μL扩增产物进行琼脂糖凝胶电泳,确定扩增片段的大小符合要求。用试剂盒提供的纯化柱进行纯化去掉200 bp以下的小片段。电泳结果显示纯化后的双链cDNA的大小主要在300~
3 000 bp之间(图2)。
2.3文库的容量与滴度
根据公式计算得知9311的初始文库独立克隆子数目为2.22×106 CFU;用液体培养基收集后,血球计数板检测文库的滴度为5.4×107 CFU/mL。20多倍的文库滴度既保证了cDNA文库的完整性和覆盖度,也减少了筛库的工作量。
2.4酵母文库插入片段大小
随机挑选24个文库转化子,通过PCR反应检测文库插入片段大小。从图3中可以看出:所构建的9311酵母文库中,插入片段大小不等,具有良好的多样性;插入片段大小主要分布在400~1 500 bp之间,密集分布在700~1 000 bp之间,文库插入片段平均大小约为880 bp。
3讨论
用于构建水稻酵母双杂交文库的水稻品种是9311,其并不具有对褐飞虱的抗性。但是有研究表明通过分子标记辅助选择将抗褐飞虱基因导入9311后,育成的材料具有稳定的抗性[10,11]。考虑到9311具有优良的农艺性状,已测序完成,有可用的基因组序列,且多个抗褐飞虱基因均能在9311中发挥作用。所以选择用受褐飞虱取食诱导的9311来构建酵母双杂交文库,以用于筛选与抗性相关基因互作的蛋白。
对文库质量的评价主要体现在两个方面。一方面为文库的代表性,指文库中包含的重组cDNA分子反映来源组织中表达信息(即mRNA种类)的完整性,其好坏可用文库的库容量来表示,指构建的初始cDNA文库中所包含的独立的重组子克隆数。库容量取决于来源组织中所表达的mRNA种类和每种mRNA的拷贝数。因此,RNA的提取是cDNA合成和文库构建过程中最为关键的一步。本研究严格按照试剂盒说明抽提并纯化mRNA,通过质量评价确定所提取的RNA产量高、纯度好,未发生降解,且调整浓度至所有样品量一致。测定酵母文库的库容量大于1×106 CFU,保证了文库的完整性和覆盖度。文库质量评价的另一个方面是重组cDNA片段的序列完整性。本研究在合成cDNA的过程中,尽可能保证cDNA序列的完整性。对构建的文库cDNA插入片段PCR结果分析表明,插入片段大小主要分布在400~1 500 bp之间,密集分布在700~
1 000 bp之间,文库插入片段平均大小约为880 bp。本研究所构建的水稻品种酵母双杂交文库经分析评价确定其质量达到了酵母双杂交研究的标准,可以用来进行大规模的互作蛋白筛选实验。
在植物与病原菌相互作用的研究中,人们通过酵母双杂交系统,在不同的植物中筛选到了与抗病蛋白相互作用的蛋白分子。Mackey等[12]通过筛选拟南芥的酵母双杂交文库,发现拟南芥的抗病蛋白RPM1和RPS2通过RIN4蛋白与三种不同的病原菌Avr蛋白相互作用,使拟南芥产生抗病反应。Yuan等[13]在水稻中筛选出了与抗白叶枯基因Xa13互作的蛋白COPT1和COPT5,这两个蛋白调节水稻体内铜离子的含量,从而使水稻产生抗病反应。但是,在水稻抗虫研究中,由于并没有克隆出水稻的抗虫基因,所以很少有通过酵母双杂交系统研究水稻与昆虫相互作用的报道。目前,水稻抗褐飞虱基因Bph14已经被成功克隆出来[5],因此构建受褐飞虱取食的水稻酵母双杂交文库,能为筛选与Bph14互作的蛋白分子,从而研究水稻抗褐飞虱的分子机理奠定基础,并对研究植物与刺吸式口器昆虫的相互作用具有参考意义。
参考文献:
[1] NORMILE D. Reinventing rice to feed the world[J]. Science, 2008,321(18):330-333.
[2] TANAKA K,ENDO S,KAZANO H. Toxicity of insecticides to predators of rice planthoppers: Spiders, the mirid bug and the dryinid wasp[J]. Applied Entomology and Zoology,2000,35(1):177-187.
[3] ZHANG Q. Strategies for developing green super rice[J]. Proceedings of the National Academy of Sciences,2007,104(42): 16402-16409.
[4] RAHMAN M L, JIANG W, CHU S H, et al. High-resolution mapping of two rice brown planthopper resistance genes, Bph20(t) and Bph21(t), originating from Oryza minuta[J]. Theoretical and Applied Genetics,2009,119(7):1237-1246.
[5] DU B,ZHANG W,LIU B,et al. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice[J]. Proceedings of the National Academy of Sciences,2009,106(52):22163-22168.
[6] ZHANG F,ZHU L,HE G. Differential gene expression in response to brown planthopper feeding in rice[J]. Journal of Plant Physiology,2004,161(1):53-62.
[7] YUAN H,CHEN X,ZHU L,et al. Identification of genes responsive to brown planthopper Nilaparvata lugens St?覽l (Homoptera:Delphacidae) feeding in rice[J]. Planta,2005,221(1): 105-112.
[8] WANG Y,WANG X,YUAN H,et al. Responses of two contrasting genotypes of rice to brown planthopper[J]. Molecular Plant-Microbe Interactions,2008,21(1):122-132.
[9] WEI Z,HU W,LIN Q,et al. Understanding rice plant resistance to the brown planthopper(Nilaparvata lugens): A proteomic approach[J]. Proteomics,2009,9(10):2798-2808.
[10] 李进波,夏明元,戚华雄,等. 水稻抗褐飞虱基因Bph14和Bph15的分子标记辅助选择[J].中国农业科学,2006,39(10):2132-2137.
[11] QIU Y,GUO J,JING S,et al. High-resolution mapping of the brown planthopper resistance gene Bph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isogenic backgrounds[J]. Theoretical and Applied Genetics,2010,121(8):1601-1611.
[12] MACKEY D,HOLT B F,WIIG A. RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis[J]. Cell,2002, 108(6):743-754.
[13] YUAN M,CHUI Z,LI X,et al. The bacterial pathogen Xanthomonas oryzae overcome rice defense by regulating host copper redistribution[J]. Plant Cell,2010,22(9):3164-3176.
