HIV-1整合酶结构的研究进展
2011-11-18张智锦胡建平
张智锦 胡建平


摘要:完整而确定的HIV-1整合酶结构在基于受体的抗AIDS药物设计中起着重要的作用。概述了HIV-1整合酶的结构和功能,并从片段结构、模建的全长聚集体以及同源体系3个方面综述了整合酶结构的研究进展。给出了各个代表性体系的应用价值。旨在对HIV-1整合酶抑制剂药物设计及筛选平台的建立提供一定的参考。
关键词:HIV-1;整合酶;结构;药物设计
中图分类号:Q814;O641文献标识码:A文章编号:0439-8114(2011)15-3032-06
Research Advances on the Structure of HIV-1 Integrase
ZHANG Zhi-jin,HU Jian-ping
(Institution of Biodiversity Conservation and Utilization in Mount Emei, Life and Chemistry College, Leshan Normal University,
Leshan 614004, Sichuan, China)
Abstract: The full length of determined HIV-1 (Human immunodeficiency virus type 1) integrase plays key roles in the anti-AIDS drug design based on the structure of acceptor. The structure and function of HIV-1 integrase and the advances in three aspects, segment structures, full length of constructed multimer and homologic systems, were summarized. The application value of each representative system was evaluated to provide some guidance for the platform established for the drug design and screening of HIV-1 integrase inhibitors.
Key words: HIV-1; integrase; structure; drug design
自从1981年6月发现首例艾滋病患者以来,艾滋病在全球范围内迅速传播。艾滋病又称获得性免疫缺陷综合症(Acquired immunodeficiency syndrome,AIDS)。中国大陆在1985年发现首例AIDS,AIDS在我国的流行经历了传入期、播散期,目前正处于高速发展期。根据最近世界卫生组织(World Health Organization,WHO)和联合国艾滋病规划署(UNAIDS,http://www.unaids.org)的估算,截至2009年底,全球AIDS感染者已达3 500万人,其中成年人超过3 000万人,妇女和儿童超过1 900万人。我国AIDS病毒感染者总数已接近100万人,并呈上升趋势,若不能有效控制,今后10年这一数字可能会成倍增长,形势十分严峻。AIDS已成为威胁人类健康的大敌,对社会将造成巨大破坏。
I型人类免疫缺陷病毒(Human immunodeficiency virus type 1,HIV-1)是引起AIDS的病源。患者体内HIV-1致死CD4+细胞,干扰其正常功能,使患者免疫功能减弱,进而引发AIDS。当前AIDS治疗的主要策略是联合使用逆转录酶抑制剂和蛋白酶抑制剂的高效抗逆转录病毒疗法(Highly active anti-retroviral therapy, HAART),又称“鸡尾酒疗法”。尽管该策略在AIDS治疗上发挥了重要作用,但两类抑制剂存在较大的抗药性、高毒性及高价格。临床上迫切需要寻找新的抗AIDS药物靶点[1,2]。HIV-1整合酶(Integrase, IN)能介导逆转录病毒DNA整合到宿主细胞DNA中,整合过程包括两步反应[3]: 第一步是3端加工(3-end processing, 3EP),即IN切割并结合病毒DNA形成IN-病毒DNA复合物;第二步是链转移(Strand transfer, ST),即IN-病毒DNA复合物切割并结合宿主细胞DNA,将病毒DNA整合到宿主DNA中,并随着宿主DNA的复制而复制。如果能抑制病毒DNA的整合过程,就防止了细胞被永久性的感染,而且人体细胞中没有IN的功能类似物,使得IN抑制剂选择性很高,对于正常细胞毒性小,所以针对IN催化反应的抑制剂设计已经成为国际上抗AIDS药物研究的热点[4-6]。2007年 Raltegravir(MK20518)被批准为第一个抑制IN的新药,IN抑制剂的研究取得了重大突破[7,8]。
HIV-1 IN含288个氨基酸残基,大小为32 kD,由N-端结构域(N-terminal domain, NTD)催化核心结构域(Catalytic core domain,CCD)和C-端结构域(C-teminal domain, CTD)折叠而成。NTD是由1~49位氨基酸残基组成的螺旋-转角-螺旋结构,内含一个不保守的HHCC锌指结构,功能上主要是促进酶的四聚化和增强酶的催化活性[9]。CCD由50~212位氨基酸残基组成,含有混合的α/β结构(共5个β折叠和6个α螺旋),是IN参与催化反应的主要区域,含有核酸内切酶和聚核苷酸转移酶的酶切位点。CCD结构上有高度保守的DDE酸性基序与二价金属离子(Mn2+或Mg2+),二者螯合共同构成IN的活性中心。值得一提的是,这3个酸性关键残基分别分布在不同的结构段上,即β1无规卷曲和α4段。另外,残基范围140~149的二级结构是一个较长的无规卷曲,该区域结构的充分柔性对于IN的催化是必须的[10]。CTD由213~288位氨基酸残基组成,包含了5个反平行的β折叠,形成了一个β桶,在IN的3个结构域中保守性最小,功能上是参与结合非特异性DNA,稳定IN-DNA复合物结构[11]。一般认为,单独的CCD就能进行简单的去整合反应,但是要完全实现3EP和ST反应,3个结构域缺一不可[12]。
近年来,HIV-1 IN结构的研究是一个热点问题,引起了很多研究者的重视。有生物学家已经从蛋白质结晶以及突变试验等方面对IN的结构及催化功能进行了较为全面的研究,并取得了一定的成果。分子模拟理论工作者则主要用结构模建、分子对接以及虚拟筛选等方面进行了一些较为系统的研究。笔者从片段结构、聚集体模建和同源复合物体系的角度出发,将HIV-1 IN结构的最新研究进展进行了综述, 旨在为有效设计HIV-1 IN抑制剂提供参考。
1整合酶的片段结构
由于HIV-1 IN溶解性较差,难以稳定结晶,迄今生物学家用X-ray和NMR试验方法仍未获得完整的IN单体结构,更不用说二聚体和四聚体。但蛋白质数据库(Protein data bank, PDB)中目前已经有超过25个HIV-1 IN的片段结构。这里将综述比较有代表性的HIV-1 IN片段结构,这些结构虽然仅是IN的片段结构,但是它们对于早期的科研以及全长IN模建中起着重要的作用[13,14]。
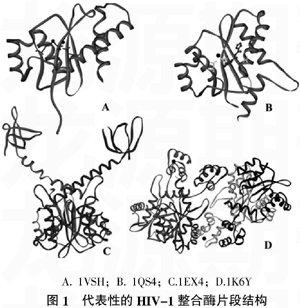
前面提到,作为聚核苷酸转移酶,HIV-1 IN含有2个金属结合区域,即结合Zn2+的NTD锌指结构和结合Mg2+或Mn2+的CCD,这些金属离子是酶催化所必须的。Bujacz等[15]用X-ray试验首次获得了含双Mg2+的CCD晶体结构(pdb代码:1VSH)。图1(A)给出了1VSH三维结构,其中用绿色Solid ribbon模式表示蛋白质部分,金属离子用黑色CPK模式表达,而DDE基序(Asp64,Asp121和Glu157)残基用粉红色Stick表示。该结构的解析首先证明了IN催化中需要双金属参与的观点一致,另外,Mg2+、Mn2+、Zn2+、Ca2+以及Cd2+均可以结合到CCD活性口袋区中。试验还表明,功能Loop区(残基138~149)的柔性是整合酶维持高活性所必须的。由于有Mg2+位置的确定,将为后续的结构解析提供了较好的参照作用。Goldgur等[16]通过双突变F185K和W131E提高IN的溶解度,并把双突变IN在大肠杆菌中表达出来,然后用X-ray方法获得了到目前为止惟一的一个IN与其抑制剂1-(5-氯-吲哚-3-甲酸)-3-四唑基-1,3-丙二酮烯醇(5CITEP)的复合物结构(pdb代码:1QS4),分辨率是2.1?魡。值得一提的是,IN部分由3个CCD组成。图1(B)给出了1QS4中一个单体的三维结构,其中蛋白质和金属离子表示方法同图1(A),而DDE基序(Asp64, Asp116和Glu152)残基和5CITEP抑制剂分别用黄色Stick以及粉红色Ball-and-stick表示。1QS4中Mg2+、DDE基序和抑制剂5CITEP高分辨率的结合模式,为后续基于IN结构的药物分子设计提供了有用结构信息。
多位点突变策略较大地提高了HIV-1 IN的溶解性,解析所得片段也逐渐变长。Chen等[17]通过引入五点突变(C56S/W131D/F139D/F185K/C180S),首次解析出包含有CCD和CTD区域的HIV-1 IN片段的对称二聚体,分子外形是一个“Y”字形,残基范围是52~288,分辨率为2.8?魡(pdb代码:1EX4),结构详见图1(C)。CCD和CTD之间靠27个氨基酸长度的α6螺旋(残基195~211)连接。可以推测,两个较长的α6柔性臂可能在整合催化中起着结构域重定向作用。值得一提的是,该结构是首次报道的多结构域IN体系,从结构上解释了IN四聚体存在的合理性。两个CCD间存在1个二体接触面,而两个CTD之间距离大约为55?魡。试验表明,CTD有专一性结合DNA的特性,推测CTD在整合催化作用中,应该有参与结合、弯转甚至重定向病毒DNA的功能。电势计算结果表明,IN二聚体中有一条沿B链CCD到A链CTD的狭长正电区域带,病毒DNA可能结合到该区域。参与结合的氨基酸残基有B链的Lys159、Lys186、Arg187和Lys188以及A链的Lys211、Lys215、Lys219、Lys243、Arg263和Arg264,这点证明了结构完整性是IN发挥其催化功能的基础。另外该课题组还解析一个单独CCD(残基范围52~212)结构,与1EX4的CCD部分完全吻合,进一步证明了1EX4结构的合理性。Wang等[18]解析出了包含有NTD和CCD区域的HIV-1 IN片段结构(pdb代码:1K6Y),形成了1对非对称的IN四聚体,CCD和CTD之间是靠一个高度无序链47~55连接,残基范围是1~212,结构详见图1(D)。该结构给出了IN四聚体中NTD以及CCD的接触界面,是模建IN四聚体的基本模板,具体做法是与包含有CCD和CTD区域的片段叠合,结构衍生模建出全长的HIV-1 IN二聚体和四聚体。为了提高溶解性以利于结晶,体系进行了三位点突变(W131D/F139D/F185K)。结构分析表明,1K6Y每个CCD或者CTD与之前报道的单体三维结构十分吻合,而且CCD与1EX4的对应区域结构类似。
2完整聚集体模建研究
体内及体外的一系列生物学试验研究[19]表明,有活性的HIV-1 IN是多聚体,而且病毒中也存在一些低聚体IN,推测IN在多聚体和低聚体之间有一个动态平衡。但是IN活性多聚体亚基数目有待深入研究[20]。基于PDB库中已有的IN片段结构,国际上已有很多小组用同源模建方法获得了全长IN二聚体和四聚体等结构。
Luca等[21]首次用同源模建方法搭建了一条完整的HIV-1 IN二聚体结构,并用ESCHER分子对接程序与一条含有27 bp的病毒DNA进行复合物预测,获得了IN与病毒DNA的结合模式(pdb代码:1WKN)。图2(A)给出了1WKN的三维结构,绿色及紫色Solid ribbon模式表示IN的两个单体,而病毒DNA用黄色Ring表示。模建过程有5个步骤:①保留1QS4 晶体结构[16]中A、B链的CCD,去除其中的配体、结晶水及C链,并把4 个缺失残基Ile141、Pro142、Tyr143和Asn144 依据1BIS[22]中B链的同源区域补全;②把晶体结构中两个突变残基F158K和W131E替换为天然残基,得到的野生型CCD二聚体的A、B链分别包含一个Mg2+;③按照包含双金属离子与HIV-1 IN高度同源的鸟肉瘤病毒IN结构1VSH[15],将第二个Mg2+放置于A、B链相应的位置;④把双金属IN2核心区与PDB代码为1EX4[17]和1K6Y[18]的结构依次叠落以获得全长的IN2;⑤最后与1WJD[23]结构叠落补全缺失的47~55 号残基。通过详细的接触残基分析,证明了搭建的全长IN二聚体-病毒DNA复合物准确验证了一系列交叉偶联和突变试验。Luca采用的病毒DNA是已经切除3末端2个核苷酸的。实际上,在生理状态下,病毒DNA先与IN结合后再发生3EP反应,随之IN和病毒DNA的构象将发生一定变化。所以3EP反应前后的结合模式可能会有所不同,基于此,有学者也用Jackal程序搭建了全长的IN二聚体[24-26]。并用Autodock程序获得了与8 bp以及27 bp病毒DNA(没有切除CA碱基)的复合物结构,发现随着病毒DNA长度的增加,HIV-1 IN2与病毒DNA的具有生物活性的结合模式出现的可能性相应增大。并用分子动力学方法研究了复合物在水溶液中的构象变化。结果表明,与未结合IN的病毒DNA相比,复合物中病毒DNA的结合区碱基构象变化较大,尤其是结合部位的小沟变宽。
最近,Wielens等[27]以及美国国家癌症研究中心Karki等[28]分别用同源模建的方法获得了全长的整合酶四聚体,并基于四聚体结构研究了整合酶与病毒DNA可能的结合模式以及金属离子对结合的影响。其中Karki等[28]提供了3个模建路径,得到3个模建结构。模型1路径具体为:①保留1EX4(CTD+CCD)二体结构,去除水,用Sybyl补全缺失残基,而功能Loop区(残基138~149)用PDB库中1BL3结构[29]补全,CTD的部分走向用PDB库中1IHV结构[30]校准;②用PDB库中1WJA结构[23]补全NTD部分,这样得到了全长IN二聚体;③与1K6Y四聚体[18]叠落,获得全长四聚体;④放置两个金属Mg2+在DDE口袋中,第一个手工放置,第二个参照1QS4结构[16]。图2(B)给出了模型1结构。模型2的路径与模型1的区别有两点:①NTD结构采用1K6Y结构[18]而不是1WJA[23];②两个Mg2+的位置参照1VSH结构[15],而非手动以及参照1QS4[16]。图2(C)给出了模型2结构。模型3的路径与模型2相近,只是模型3中CTD的部分走向未用PDB库中1IHV结构[30]校准,图2(D)给出了模型3结构。Karki等[28]的研究成果首次给出了IN与病毒DNA及宿主DNA的识别结果,证明Glu152、Gln148和Lys156是结合病毒DNA的反应部位(CA-OH3)的关键残基;以及功能Loop区(残基140~149)主要是通过隔开病毒DNA和宿主DNA,从而起到稳定IN和复合物的作用。Wielens等[27]还用GRID程序将一系列IN抑制剂对接到IN四聚体-病毒DNA-宿主DNA上,这些对接结果都与已有的试验信息相吻合,并且对IN与DNA结合模式提供了一些有用的信息。Wang等[31]搭建了一个与Karki等[28]提供的类似全长的IN四聚体结构,首次定量考虑了金属离子对于IN与病毒DNA相互作用的影响;Ke等[32]利用PDB库中已有的片段结构,也搭建了一个IN四聚体,并把不同长度的病毒DNA对接到该平台中,结果表明,IN四聚体中有3个DNA结合区域,有两个应该是病毒DNA结合区,另一个是宿主DNA结合区。
3同源蛋白酶-DNA复合物结构
在HIV-1的生命周期中,HIV-1 IN主要是将病毒DNA整合到宿主细胞染色体中,整个催化反应前提是IN与病毒DNA的特异性识别与结合。在生理状态下,病毒DNA先与IN结合,随后发生3EP反应。在药物设计中,如果知道HIV-1 IN与病毒DNA的结合位点,就可以有针对性地寻找阻断HIV-1 IN与病毒DNA结合的抑制剂,阻断HIV-1的复制过程。尽管上面提到了Luca等[21]、Wielens等[27]和Karki等[28]均搭建出IN-DNA模型,这些为HIV IN的催化机理以及IN抑制剂的合理设计研究提供了许多有益的信息,但是它们依然无法完全满足当前IN抑制剂设计工作的需要。因为在以往的IN-DNA模型构建过程中,涉及到IN二聚体、四聚体以及病毒DNA的模建,模建误差对后续计算结果准确性会造成一定的影响。总之,无论单独的全长IN还是与病毒DNA的复合物结构都缺失,尽管HIV整合酶抑制剂Raltegravir已经用于临床,但是其作用机制仍未知。
Steiniger等[33]解析出Tn5转座酶与DNA末端形成的复合物结构(pdb代码:1MUS),首次提供了转座/整合路径分析的三维结构。细菌Tn5转座酶和HIV-1 IN同属于聚核苷酸转移酶家族,催化机制类似。Tn5转座酶常用于DNA转移研究的模型系统,和HIV-1 IN一样内含3个保守的氨基酸残基(Asp97、Asp188、Glu326,即DDE基序),与金属Mn2+稳定结合,共同对酶的DNA转移活性起关键性的作用。Tn5-DNA复合物结构中的Mn2+被Asp97、Glu326和DNA的3′-OH所定位,共同构成的活性口袋在三维结构上与HIV-1 IN非常相似。与HIV-1
IN一样,Tn5转座酶的活性口袋区中含多个Lys残基,参与结合DNA末端序列。另外,Tn5活性口袋区中的两个金属离子得到解析,支持双金属参与IN催化的观点。在HIV-1 IN-DNA复合物结构缺乏的情况下,该结构是一个很好开展DNA转移机制研究的平台[34]。图3(A)给出了Tn5转座酶的三维结构,两个Mn2+和DDE基序残基分别用黑色CPK和粉红色Stick表示,蛋白质和DNA部分则分别用绿色Solid ribbon和黄色Stick表示。
最近,Hare等[35]用X-ray方法解析出全长的原核泡沫病毒(Prototype foamy virus,PFV)IN与病毒DNA的复合体三维结构(pdb代码:3OYA)。晶体结构由PFV IN四聚体和两条病毒DNA组成,其中PFV IN与HIV-1 IN高度同源,均属于聚核苷酸转移酶家族。PFV IN保留了HIV-1 IN中高度保守的DDE活性口袋区以及双金属催化中心。Tang等[36]的对接试验表明,一系列HIV-1 IN抑制剂在PFV IN中的结合位置和在HIV-1中的结合位置相同,均是在DDE活性口袋区。鉴于晶体库中PFV IN含HIV-1 IN没有但又十分重要的一些结构信息,比如酶多聚化和病毒DNA的结合位置等,所以PFV IN可以作为基于结构的抗AIDS药物研发的一个有效靶点。
4结语
从原子水平上研究HIV-1 IN催化位点的结构信息和作用机理,进而为基于结构的IN抑制剂设计具有重要的意义。本文把握国内外当前对IN结构研究的前沿,总结了该领域一系列代表性成果,希望借此引起读者的关注。
参考文献:
[1] PERRYMAN A L, FORLI S, MORRIS G M, et al. A dynamic model of HIV integrase inhibition and drug resistance[J]. Journal of Molecular Biology,2010,392(2):600-615.
[2] KOTOVA S, LI M, DIMITRIADIS E K, et al. Nucleoprotein intermediates in HIV-1 DNA integration visualized by atomic force microscopy[J]. Journal of Molecular Biology,2010,399(3):491-500.
[3] DOLAN J,CHEN A P,WEBER I T,et al. Defining the DNA substrate binding sites on HIV-1 integrase[J]. Journal of Molecular Biology,2009,385(2):568-579.
[4] MCCOLL D J, CHEN X W. Strand transfer inhibitors of HIV-1 integrase: Bringing IN a new era of antiretroviral therapy[J]. Antiviral Research,2010,85(1):101-118.
[5] 胡建平,唐典勇,范晶,等. HIV-1整合酶G140S/G149A及T66I/S153Y突变后的构象变化[J]. 化学学报,2010,68(15):1499-1506.
[6] HU J P, GONG X Q, SU J G, et al. Study on the molecular mechanism of inhibiting HIV-1 integrase by EBR28 peptide via molecular modeling approach[J]. Biophysical Chemistry, 2008,132(2):69-80.
[7] SUMMA V, PETROCCHI A, BONELLI F, et al. Discovery of raltegravir, a potent, selective orally bioavailable HIV-integrase inhibitor for the treatment of HIV-AIDS infection[J]. Journal of Medicinal Chemistry,2008,51(18):5843-5855.
[8] ALIAN A, GRINER S L, CHIANG V, et al. Catalytically-active complex of HIV-1 integrase with a viral DNA substrate binds anti-integrase drugs[J]. Proceedings of the National Academy of Sciences USA,2009,106(20):8192-8197.
[9] HEUER T S, BROWN P O. Photo-cross-linking studies suggest a model for the architecture of-118-An active human immunodeficiency virus type 1 Integrase-DNA Complex[J]. Biochemistry,1998,37:6667-6678.
