AD5-10与表阿霉素联合作用治疗类风湿性关节炎
2011-11-16周剑锁朱洁卿袁海芹刘彦信郑德先
周剑锁,史 娟,朱洁卿,袁海芹,刘彦信,尤 欣,郑德先
中国医学科学院 北京协和医学院 基础医学研究所医学分子生物学国家重点实验室,北京100005
AD5-10是本实验室此前发现并报道的针对人死亡受体5(death receptor,DR5)的单克隆抗体,在体内外对多种肿瘤细胞系有抗肿瘤作用,而对正常细胞没有毒性。在一定条件下,AD5-10可激活caspase依赖型和caspase非依赖型两种细胞死亡信号通路[1-2]。其与 DR5的结合位点位于 DR5的 N末端[3]。敲除c-FLIPL可以加强肺癌细胞对 AD5-10的敏感性[4],提示c-FLIPL可能在AD5-10的肿瘤治疗及某些AD5-10抗性细胞杀伤中发挥作用。
表阿霉素 (epirubicin,EPB)是一种蒽环类抗生素,为阿霉素的同分异构体,毒性较阿霉素低,是当前治疗多种癌症的临床一线用药[5-7]。
类风湿性关节炎 (rheumatoid arthritis,RA)是一种以关节滑膜炎症为特征的慢性全身性自身免疫性疾病。滑膜炎是RA的基本病理改变,主要表现为滑膜炎性细胞浸润和血管增生以及软骨和软骨下骨组织破坏,RA滑膜表面的成纤维样滑膜细胞 (fibroblast-like synoviocytes,FLS)扩张性增殖。引起RA关节滑膜组织过度增生的基本原因可能是滑膜细胞凋亡机制障碍导致的滑膜组织内细胞增殖和死亡间的平衡失调。有研究发现,虽然FLS表面表达一些凋亡受体,但其净效应是细胞增殖而非细胞凋亡[8]。相当一部分学者认为,FLS在RA的长期病程中起到关键作用[9]。诱导FLS发生程序性细胞死亡是RA治疗的一个重要策略。本研究观察了AD5-10和表阿霉素联合作用治疗RA的分子机制。
材料和方法
试剂AD5-10单克隆抗体为本实验室制备[1]。抗caspase-8、-9、-3,Bcl-2和 cIAP1抗体购自美国CST公司,抗p53和p21抗体购自美国Santa Cruz公司,抗c-FLIP抗体购自美国Alexis公司,辣根过氧化物酶标记的二抗混合物购自中杉金桥公司。
滑膜细胞分离培养实验所用FLS细胞从RA患者膝关节置换术后关节切除物分离,由北京协和医学院风湿免疫科提供。将滑膜组织剪成1 mm2大小碎块,加入1 mg/mlⅠ型胶原酶 (美国Sigma公司)处理2 h,放入37℃含5%CO2的孵箱中培养24~48 h,倒去未贴壁的细胞,剩余贴壁细胞在含有10% 小牛血清(美国Hyclone公司)、100 U/L青霉素及100 mg/L链霉素的100 ml DMEM培养基 (美国Invitrogen公司)中继续培养,每3~4 d换液1次。用于实验的细胞均为5~10代传代细胞。
MTT法测定细胞活性收集对数期细胞,调整细胞悬液浓度至105/ml,加入96孔细胞培养板,每孔100 μl,在5%CO2和37℃条件下孵育至细胞单层铺满孔底,用不同浓度AD5-10(0、10、50、250和1250 ng/ml)单独处理FLS,或以上述不同浓度AD5-10配合不同浓度EPB(50、200和1000 ng/ml)联合用药处理。加入药物后继续孵育12 h。每孔加入20 μl MTT溶液 (终浓度1 mg/ml),继续培养4 h。小心吸去孔内培养液,每孔加入150 μl二甲基亚砜,置摇床上低速振荡10 min,使结晶物充分溶解。在酶联免疫检测仪570 nm处测量各孔的吸光值 (optical density,OD)。取3个平行孔的平均值,按公式计算细胞存活率 (%):加药组OD值/对照组OD值×100%。
免疫印迹以蛋白裂解液[1%NP-40、0.7 mmol/L EDTA、1∶9(v/v)蛋白酶抑制剂]裂解细胞,包括未经药物处理、400 ng/ml EPB单独处理、500 ng/ml AD5-10单独处理,及EPB和AD5-10联合处理的细胞。SDS聚丙烯酰胺凝胶电泳分离蛋白,电转PVDF膜后,行蛋白免疫印迹。转膜成功后,5%脱脂奶粉封闭1 h,TBST洗膜后加一抗,4℃孵育过夜,TBST洗膜。加辣根过氧化物酶标记二抗,室温孵育2 h,TBST洗膜,将化学发光试剂ECL(美国GE&Millipore公司)加至膜上,最后X片显影。
结 果
表阿霉素提高FLS对AD5-10的敏感性以MTT法测定经不同浓度AD5-10处理的FLS细胞的存活率,结果显示AD5-10的浓度达到1250 ng/ml时细胞存活率仍无变化 (图1)。加入不同浓度EPB联合处理时,FLS细胞存活率降低,且呈浓度依赖性 (图2)。
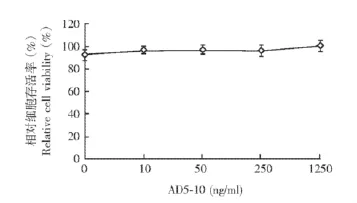
图1 成纤维样滑膜细胞对AD5-10不敏感Fig 1 Fibroblast-like synoviocytes show resistance to AD5-10
EPB和AD5-10联合作用对细胞凋亡信号通路的影响用AD5-10(500 ng/ml)或EPB(400 ng/ml)单独处理时,细胞中的procaspase-3、procaspase-8和procaspase-9的水平没有显著变化,但联用时3种酶原明显被激活 (图3)。
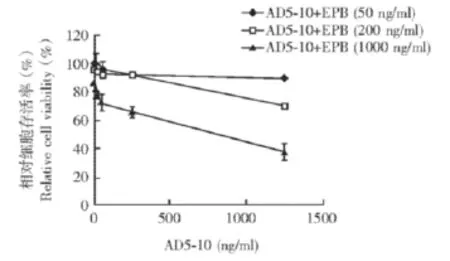
图2 表阿霉素可提高成纤维样滑膜细胞对AD5-10的敏感性Fig 2 Epirubicin enhances the sensibility of fibroblast-like synoviocytes to AD5-10
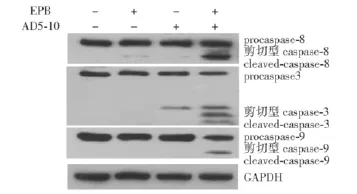
图3 AD5-10(500 ng/ml)和表阿霉素 (400 ng/ml)单独应用和联合应用时,细胞中procaspase-3、procasepase-8和procaspase-9的激活情况Fig 3 The activation of procaspase-3,procaspase-8,and procaspase-9 when treated with AD5-10(500 ng/ml),epirubicin(400 ng/ml),or the two agents in combination
EPB增强FLS对AD5-10敏感性的分子机制EPB单独处理对凋亡抑制蛋白Bcl-2和cIAP1的表达没有影响,但可显著下调caspase-8竞争结合蛋白c-FLIP的表达水平,而EPB与AD5-10联合处理组的Bcl-2水平有所下调。EPB可增强p53表达,抑制p21表达 (图4)。
讨 论
RA是一种复杂的多系统疾病,其炎症性组织损伤的初始部位是关节。研究显示,FLS在关节损伤和炎症进展方面发挥重要作用[9]。诱导滑膜衬里层细胞发生程序性细胞死亡是RA治疗的一个重要策略。其中,可以特异性诱导肿瘤细胞发生凋亡的TNF家族成员TRAIL也是RA治疗研究的热点,但TRAIL或DR5抗体是否可引起RA-FLS凋亡还存在争议。有研究发现TRAIL或DR5的抗体可诱导FLS凋亡,并在RA动物模型中观察到治疗作用[10-13];但也有研究报道TRAIL会引起FLS增殖[14-15]。
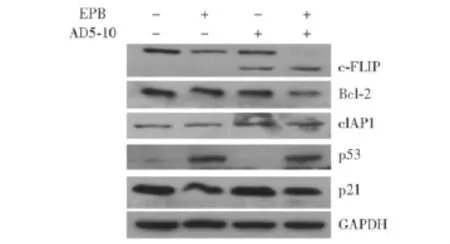
图4 AD5-10(500 ng/ml)和表阿霉素 (400 ng/ml)单独应用和联合应用时,细胞内凋亡信号通路相关蛋白及细胞周期蛋白的变化Fig 4 The changes of intracellular apoptosis pathway related proteins and cell cycle proteins after the treatment with AD5-10(500 ng/ml)alone,epirubicin(400 ng/ml)alone,or AD5-10 and EPB in combination
AD5-10是针对DR5的单克隆抗体,可以通过结合DR5,激活caspase级联和非caspase级联反应导致细胞凋亡[1]。笔者以往研究也发现AD5-10对FLS没有明显的凋亡诱导作用,因此如何提高FLS对AD5-10的敏感性是本研究的重点。与化疗药物联用是提高抗性肿瘤细胞对TRAIL或DR4/DR5的抗体敏感性的重要策略。笔者通过实验筛查多种化疗药物后,发现蒽环类抗肿瘤药物EPB与AD5-10联合作用可以很好地杀伤FLS。虽然蒽环类化疗药物未用于RA的治疗,但目前临床上已经有一些化疗药物用于治疗RA,包括甲氨蝶呤、硫唑嘌呤、环磷酰胺等。本研究结果显示不同剂量的EPB与AD5-10联用增加了FLS的凋亡。
c-FLIP是一种细胞存活信号分子,与caspase-8和caspase-10有高度同源序列,但是没有蛋白酶活性,不能发生剪切,可以干扰caspase-8结合DISC复合物中的FADD,抑制随后的caspase级联反应,从而抑制细胞凋亡[4,16]。以往研究表明,c-FLIP在RA的FLS中表达水平增高[17]。本研究发现EPB可降低c-FLIP的表达水平,这可能是EPB可以提高FLS对AD5-10敏感性的原因之一。此外,联合处理组Bcl-2的水平降低。Bcl-2有抗凋亡作用,其表达水平降低可以进一步诱导线粒体凋亡途径,使凋亡信号继续放大[18]。
本研究还发现,EPB处理后,细胞p21水平下调。p21是一种细胞周期负调控蛋白,表达水平上调可使细胞周期阻滞在G0期,下调可使细胞重新进入细胞周期,p21可抑制细胞凋亡,下调时可使细胞从生长阻滞转向凋亡[19]。FLS对 FASL和 TRAIL诱导凋亡的敏感性有细胞周期依赖性[20],因此在EPB增强FLS对AD5-10敏感性的方面,p21可能也发挥了重要作用,但EPB使p21下调的机制尚未明确。
综上,本研究结果显示,EPB可使具有抗凋亡功能的c-FLIP表达量下调,同时使细胞周期负调控蛋白p21表达量降低,显著增加FLS对AD5-10的敏感性,为RA治疗提出了新的策略。
[1]Guo Y,Chen C,Zheng Y,et al.A novel anti-human DR5 monoclonal antibody with tumoricidal activity induces caspasedependent and caspase-independent cell death [J].J Biol Chem,2005,280(51):41940-41952.
[2]Chen C,Liu Y,Zheng D.An agonistic monoclonal antibody against DR5 induces ROS production,sustained JNK activation and Endo G release in Jurkat leukemia cells[J].Cell Res,2009,19(8):984-995.
[3]Zhang P,Zheng Y,Shi J,et al.Targeting a novel N-terminal epitope of death receptor 5 triggers tumor cell death[J].J Biol Chem,2010,285(12):8953-8966.
[4]Chen F,Guo J,Zhang Y,et al.Knockdown of c-FLIPLenhanced AD5-10 anti-death receptor 5 monoclonal antibodyinduced apoptosis in human lung cancer cells[J].Cancer Sci,2009,100(5):940-947.
[5]Delaney CE,Hopkins SP,Addison CL.Supplementation with L-carnitine does not reduce the efficacy of epirubicin treatment in breast cancer cells [J].Cancer Lett,2007,252(2):195-207.
[6]周佳彦,段金虹,孙昭,等.白藜芦醇与表阿霉素联合用药对多种人类肿瘤细胞的体外抑制作用[J].基础医学与临床,2010,30(11):1149-1152.
[7]Plosker GL,Faulds D.Epirubicin,a review of its pharmacodynamic and pharmacokinetic properties,and the rapeutic use in cancer chemotherapy[J].Drugs,1993,45(5):788-856.
[8]Palao G,Santiago B,Galindo M,et al.Down-regulation of FLIP sensitizes rheumatoid synovial fibroblasts to Fas-mediated apoptosis[J].Arthritis Rheum,2004,50(9):2803-2810.
[9]周振华,徐卫东,吴岳嵩.滑膜成纤维细胞与类风湿性关节炎相关研究的进展[J].中华关节外科杂志,2008,2(6):692-696.
[10]Song K,Chen Y,Göke R,et al.Tumor necrosis factor-related apoptosis-inducing ligand(TRAIL)is an inhibitor of autoimmune inflammation and cell cycle progression[J].J Exp Med,2000,191(7):1095-1104.
[11]Ichikawa K,Liu W,Fleck M,et al.TRAIL-R2(DR5)mediates apoptosis of synovial fibroblasts in rheumatoid arthritis[J].J Immunol,2003,171(2):1061-1069.
[12]Yao Q,Seol DW,Mi Z,et al.Intra-articular injection of recombinant TRAIL induces synovial apoptosis and reduces inflammation in a rabbit knee model of arthritis[J].Arthritis Res Ther,2006,8(1):R16.
[13]Terzioglu E,Bisgin A,Sanlioglu AD,et al.Concurrent gene therapy strategies effectively destroy synoviocytes of patients with rheumatoid arthritis [J].Rheumatology(Oxford),2007,46(5):783-789.
[14]Morel J,Audo R,Hahne M,et al.Tumor necrosis factorrelated apoptosis-inducing ligand(TRAIL)induces rheumatoid arthritis synovial fibroblast proliferation through mitogenactivated protein kinases and phosphatidylinositol 3-kinase/Akt[J].J Biol Chem,2005,280(16):15709-15718.
[15]Audo R,Combe B,Coulet B,et al.The pleiotropic effect of TRAIL on tumor-like synovial fibroblasts from rheumatoid arthritis patients is mediated by caspases[J].Cell Death Differ,2009,16(9):1227-1237.
[16]Krueger A,Baumann S,Krammer PH,et al.FLICE-inhibitory proteins:regulators of death receptor-mediated apoptosis[J].Mol Cell Biol,2001,21(24):8247-8254.
[17]Schedel J,Gay RE,Kuenzler P,et al.FLICE-inhibitory protein expression in synovial fibroblasts and at sites of cartilage and bone erosion in rheumatoid arthritis[J].Arthritis Rheum,2002,46(6):1512-1518.
[18]Cheng EH,Wei MC,Weiler S,et al.BCL-2,BCL-X(L)sequester BH3 domain-only molecules preventing BAX-and BAK-mediated mitochondrial apoptosis [J].Mol Cell,2001,8(3):705-711.
[19]Jung YS,Qian Y,Chen X.Examination of the expanding pathways for the regulation of p21 expression and activity[J].Cell Signal,2010,22(7):1003-1012.
[20]Pundt N,Peters MA,Wunrau C,et al.Susceptibility of rheumatoid arthritis synovial fibroblasts to FasL-and TRAIL-induced apoptosis is cell cycle-dependent[J].Arthritis Res Ther,2009,11(1):R16.
