简并PCR结合RACE技术克隆申克孢子丝菌未知过氧化氢酶基因
2011-09-11王晓慧刘伟李若瑜
王晓慧 刘伟 李若瑜
(1.厦门大学附属中山医院皮肤科,厦门 361004;2.北京大学真菌和真菌病研究中心北京大学第一医院皮肤科真菌室,北京 100034)
双相真菌申克孢子丝菌 (Sporothrix schenckii)是孢子丝菌病的致病菌,主要引起皮肤及皮下组织感染,也可以引起黏膜、骨骼或系统性病变。近年来该菌引起播散型或系统性感染增多[1-2],在免疫抑制患者尤其是艾滋病患者中发病率逐年增高[3-4],逐渐受到重视。S.schenckii致病机
制至今尚不清楚,本研究采用简并PCR联合快速cDNA末端扩增 (rapid amplification of cDNA ends,RACE)技术,成功克隆了S.schenckii过氧化氢酶(catalase,CAT)基因,命名为Sscat基因,为进一步探讨CAT在S.schenckii抗氧化防御机制中的作用奠定了基础。
1 材料与方法
1.1 实验菌株
实验菌株为S.schenckiiBMU03906,分离自皮肤淋巴管型孢子丝菌病患者,北京大学真菌和真菌病研究中心保存菌种。该菌株经双相性及菌落形态鉴定后,分别接种于马铃薯葡萄糖琼脂 (PDA)和脑心浸汁葡萄糖琼脂培养基在25℃和36℃保存。温度实验证实该菌株在37℃不生长,36℃可以生长。菌株在脑心浸汁血琼脂葡萄糖培养基36℃培养条件下,经过连续多次转种后培养出纯酵母相。实验前活化菌株。
1.2 试剂
LA Taq DNA聚合酶、T4DNA连接酶、5'RACE和3'RACE试剂盒购自TaKaRa公司;纯化mRNA用Oligo dT纤维素购自Omega公司;以pBluescript SK(+)质粒为基础构建T-克隆载体,用于克隆PCR产物。
1.3 DNA,RNA提取及cDNA合成
将实验菌株接种于PDA斜面,25℃培养7 d,转种于脑心浸汁血琼脂葡萄糖培养基,36℃培养,每7~9 d转种1次以获得酵母相。收集酵母相菌体,用氯化苄法提取 DNA[5]。Trizol法提取总RNA,经Oligo dT纤维素柱纯化mRNA,逆转录合成第一链cDNA。
1.4 简并引物设计
在NCBI GenBank中寻找白念珠菌、新生隐球菌、荚膜组织孢浆菌、巴西副球孢子菌、裂殖酵母、枝孢霉以及构巢曲霉CAT的cDNA序列及氨基酸序列,经序列比对找出上、下游两段高度保守区(见图1),设计简并引物 (见表1)。
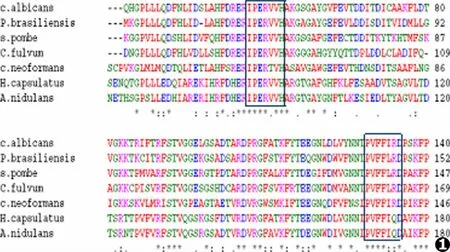
图1 7种真菌过氧化氢酶氨基酸序列的比较(方框为两个高度保守的区域)Fig.1 Alignment of catalase amino acids from seven fungal species
1.5 PCR扩增 cDNA
以酵母相S.schenckii的第一链cDNA为模板,将上、下游简并引物两两配对,采用二步PCR方法扩增。简并 PCR 反应体系:cDNA 1.0 μL,10×PCR bufferⅡ (含 Mg2+2.5 mmol/L)2.5 μL,LA Taq DNA 聚合酶 (5 U/μL)0.5 μL,上下游引物各1 μL(20 pmol/μL),dNTP 8 μL(2.5 mmol/L),加水至50.0 μL。反应条件采用两步法:94℃ 30 s,45℃ 30 s共5个循环,72℃ 30 s共5个循环,94℃30 s共5个循环94℃ 30 s共30个循环,55℃ 30 s共30个循环,72℃ 30 s共30个循环,然后72℃延伸10 min。PCR产物常规纯化、克隆、测序。所得序列和其他已知真菌CAT基因cDNA序列进行比对,证实为目的片段后进行下一步实验。

表1 简并引物及用于3'-和5'-RACE的引物Tab.1 Degenerate Primers and primers used in RACE
1.6 3'RACE 和5'RACE
根据上述获得的cDNA序列分别设计3'RACE和5'RACE引物(见表1)。3'RACE主要过程:0.2 μg mRNA经Oligo dT-3 Sites Adaptor primer反转录合成cDNA,进行PCR扩增。5'RACE主要过程:以0.1 μg mRNA 为模板,用 5'-RT-primer反转录合成cDNA第一链,RNase H降解去除DNA-RNA杂交体中的RNA,单链cDNA环化后进行巢式PCR反应。二者PCR产物常规纯化、克隆、测序。
1.7 cDNA全长克隆测序及序列分析
将上述3种方法得到的三段序列进行拼接得到Sscat基因cDNA全长序列,将cDNA及其编码氨基酸序列进行比对分析。根据Sscat基因cDNA全长的头尾序列设计1对验证引物,用cDNA作模板,扩增出Sscat基因cDNA的全长编码区,克隆测序以进一步验证。
2 结 果
2.1 简并PCR结果
PCR产物电泳显示有多个条带,但以预期大小约268 bp处条带最清晰 (见图2),对该片段切胶纯化、克隆、测序,并与其他真菌的CAT基因cDNA序列比对,证实该产物为一潜在的在Sscat基因cDNA片段。
2.2 3'RACE 及5'RACE
3'RACE扩增产物为400 bp和1 400 bp两条清晰条带,分别对以上片段进行纯化、克隆、测序及序列比对分析,证实1 343 bp产物为目的片段,400 bp为非特异扩增 (见图3)。5'RACE扩增出600 bp单一清晰条带,无其他非特异性扩增(见图4),该片段经纯化、克隆、测序及序列比对分析,证实为691 bp目的片段。
2.3 序列分析
将简并PCR产物、3'RACE及5'RACE所获序列进行对位拼接,获得一条全长为1 746 bp的序列,它与一次扩增cDNA全长所获得的序列相同。其中开放读码框 (ORF)1 500 bp,编码499个氨基酸,起始密码子ATG位于全长序列第122~124位,终止密码子 TGA位于1 619~1 621位,5'端121 bp非编码区,3'端109 bp非编码区。该序列与多个真菌的cDNA及氨基酸同源(见图5)。
2.4 序列递交NCBI Genbank(Genbank accession numberEU 364509)
2.5 内含子确定
Sscat基因的cDNA与其基因组DNA的序列比较分析,Sscat基因包含1个71 bp内含子,位于993~1 063位。
3 讨 论
目前获得全长cDNA克隆的分子生物学技术主要有3种:cDNA文库筛选法、以PCR为基础的cDNA末端快速扩增法以及基于生物信息学的电子克隆技术。RACE是1988年由Forman等[6]报道的以PCR为基础获得目的cDNA完整序列的技术,它包括5'RACE和3'RACE。5'端和3'端的克隆与序列测定是全长基因分析中的关键,因此该技术已经被广泛应用于人类、昆虫以及致病真菌未知基因克隆,如新生隐球菌、组织胞浆菌、马内菲青霉等。席丽艳应用RACE技术克隆了马内菲青霉的Ras相关基因PmRsr1[7]。曹存巍应用简并PCR结合RACE技术克隆了马内菲青霉与氧化应激相关的SKN7基因[8]。

图2 简并PCR扩增结果:1,2,5,6.简并PCR扩增产物显示多个清晰条带;M.marker 100 bp 图3 3'-RACE扩增结果 图4 5'-RACE扩增结果 图5 过氧化氢酶氨基酸序列同源性比较Fig.2 Degenerate PCR:1,2,5,6.Amplication bands;M.marker 100 bp Fig.3 Amplication of 3'-RACE Fig.4 Amplification of 5'-RACE Fig.5 Multiple alignment of the Sporothrix schenckii catalase with highly similar catalases
我们应用简并PCR结合RACE技术首次克隆了申克孢子丝菌的过氧化氢酶基因Sscat,成功的关键在于以下3方面。①引物的合理设计:简并PCR扩增未知基因片段通常由于非特异性产物而难以获得预期的目的片段,因此简并引物的设计成为简并PCR成功的先决条件。引物的简并性越低,PCR产物的特异性越强。我们分别设计了2条上游简并引物和3条下游简并引物,尽可能选择简并性小的氨基酸来设计引物,并且避免了3'末端的简并,从而降低了引物简并度,并且将上下游简并引物两两配对进行PCR扩增。②PCR扩增条件的优化:PCR扩增先在非严谨的低退火温度 (45℃)下进行5个循环,随后在较严谨的退火温度下进行25~35个循环。简并PCR应当在1~3 μmol/L比较高的引物浓度下进行,有利于目的条带的获得。③合适的模板浓度:适量cDNA模板可以减少非特异性扩增产物。此外,实验中应严格遵循RNA提取的操作规程以防RNA降解,纯化出mRNA以获得高丰度的模板,从而提高cDNA末端的扩增效率。在现阶段S.schenckii基因组测序工作尚未完成的情况下,简并PCR结合RACE技术是克隆S.schenckii未知基因的有效方法,值得推广应用。
需氧微生物具有完整的抗氧化防御系统,CAT是最重要的抗氧化酶之一,它能将H2O2分解为H2O和O2,保护微生物免受H2O2损伤。CAT分为:锰过氧化氢酶(该酶只存在原核生物中);双功能复合酶过氧化氢酶一过氧化物酶 (catalase-peroxidase,cpeA);单功能过氧化氢酶,又包括大亚基单位(80 kDa左右)和小亚基单位的过氧化氢酶(即过氧化物酶体的过氧化氢酶)。Sscat基因cDNA全长1 746 bp,包含1个内含子,其中ORF 1 500 bp,编码499个氨基酸,分子量为56.07 kDa,其氨基酸序列与其他真菌的CAT氨基酸序列具有较高的同源性,其中与米曲霉的未命名CAT同源性最高,为66.3%,与黑曲霉、棒曲霉、土曲霉的同源性分别为56.6%、55.1%和46.7%,说明真菌的 CAT 较为保守。根据Sscat编码氨基酸序列的结构特征,推测其为小亚基单位过氧化氢酶。
致病真菌侵入宿主后面临诸多环境应激,如温度变化、高渗环境、pH改变以及氧化应激。CAT是致病真菌抗氧化防御体系中的关键酶之一,可以保护真菌在宿主机体微环境中存活并且致病。在其他双相性真菌中,如马尔尼菲青霉、巴西副球孢子菌、组织胞浆菌和皮炎芽生菌,CAT均显示有抵抗氧化损伤,帮助致病菌存活并参与致病的作用[9-12]。我们研究了Sscat基因在体外双相转化时以及在H2O2应激中mRNA表达水平的差异,realtime PCR结果显示,Sscat基因的mRNA表达均增高,推测CAT在S.schenckii抗氧化损伤中发挥了作用[13]。但是,这种抗氧化作用如何参与环境应激的具体机制,尚有待于进一步研究证实。总之,Sscat基因的克隆为深入探讨S.schenckii Sscat基因结构、蛋白质表达以及基因功能奠定了基础。
致谢 北京大学第一医院中心实验室卜定方老师对本研究的技术指导。
[1]李珊山,刘鹤松,郑华,等.皮肤型孢子丝菌病585例临床分析[J].中华皮肤科杂志,2011,44(3):161-164.
[2]吴昌昊,吴旭,张立溪,等.肺部孢子丝菌病1例[J].南方医科大学学报,2009,10:2147.
[3]Callens SF,Kitetele F,Lukun P,et al.PulmonarySporothrix schenckiiinfection in a HIV positive child[J].J Trop Pediatr,2006,52:144-146.
[4]Vilela R,Souza GF,Fernandes Cota G,et al.Cutaneous and meningeal sporotrichosis in a HIV patient[J].Rev Iberoam Micol,2007,24:161-163.
[5]朱衡,瞿峰.利用氯化苄提取适于分子生物学分析的真菌DNA[J].真菌学报,1994,13:34-40.
[6]Frohman MA,Dush MK,Martin GR,et al.Rapid production of full-length cDNA,from rare transcripts:amplification using a single gene-specific oligonucleotide primer[J].Proc Nat Acad Sci USA,1998,85:8998-9002.
[7]Feng P,Xie Z,Sun J,et al.Molecular cloning,characterization and expression of PmRsr1,a Ras-related gene from yeast form ofPenicillium marneffei[J].Mol Biol Rep,2010,37(7):3533-3540.
[8]Cao C,Liu W,Li R,et al.Penicillium marneffeiSKN7,a novel gene,could complement the hypersensitivity ofS.cerevisiaeskn7 disruptant strain to oxidative stress[J].Mycopathologia,2009,168(1):23-30.
[9]Carmo JP,Dias-Melicio LA,Calvi SA,et al.TNF-alpha activates human monocytes for paracoccidioides brasiliensis killing by an H202-dependent mechanism[J].Med Mycol,2006,44(4):363-368.
[10]Howard,DH.Comparative sensitivity of histoplasma capsulatum conidiospores and bIastospores to oxidative antifungal systems[J].Infect Immun,1981,32(1):381-387.
[11]Brummer E,Sugar AM,Stevens DA.Immunological activation of polymorphonuclear neutrophils for fungal killingl studies with murine cells and Blastomyces dermatitidisin vitro[J].J Leukoc Biol,1984,36(4):505-520.
[12]张军民,章慧,孙九峰,等.过氧化氢酶家族部分成员在马尔尼菲青霉的相关研究[J].中国人兽共患病学报,2008,24(9):824-826.
[13]Wang XH,Li RY,Cao CW,et al.Differential mRNA expression ofSporothrix schenckiicatalase gene in two growth phases and effect factors[J].Chin Med J(Engl),2008,121(20):2100-2102.
