藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局
2011-08-20段敏杰高清竹郭亚奇万运帆李玉娥
段敏杰,高清竹,郭亚奇,万运帆,李玉娥,
干珠扎布1,2,旦久罗布3,韦兰亭3,西饶卓玛3
(1.中国农业科学院农业环境与可持续发展研究所,北京100081;2.农业部农业环境与气候变化重点实验室,北京100081;3.西藏自治区那曲地区草原站,西藏 那曲852100)
物种多样性是全球变化研究的重要内容之一,也是当前现代生态学和环境学研究的热点问题之一。各种各样的生物资源是地球上人类赖以生存的物质基础,不论是发展中国家还是发达国家都面临着物种多样性的保护和持续利用的问题[1]。在很大程度上,草地生态系统的稳定性和生产力的维持依赖于草地植物群落的物种多样性[2-3]。可以说,物种多样性是衡量一个地区生物资源丰富程度的重要客观指标[4-5]。物种多样性沿海拔梯度的变化规律是生物多样性研究的重要议题,海拔梯度的变化综合了温度、湿度和光照等多种环境因子,因此,在一定程度上能够显著地影响植物种群的空间分布格局[6-7]。众多研究表明,物种多样性随着海拔梯度的变化有多种形式,其中普遍认为二者呈负相关关系,即随着海拔高度的增加物种多样性降低[8-9];另一种形式是“中间高度膨胀”,即随着海拔高度的增加,物种多样性呈先增加、后减少的趋势[10-11]。
青藏高原被称为世界“第三极”,具有独特的自然地理条件,对于此研究区植物物种多样性的研究具有重要的意义。但目前针对该区域物种多样性海拔分布格局的研究不多,杨元合等[12]研究表明,青藏高原高寒草地植物物种丰富度随海拔的上升呈减少趋势,而贺连选和刘宝汉[13]对互助县的研究表明,随着海拔梯度的逐渐升高,多样性指数和均匀度增加,物种丰富度先增加后减少。藏北地区位于青藏高原腹地,地域广阔,地形复杂,气候资源呈多样性和复杂性的特点,平均海拔在4 500m以上,被称为“世界屋脊”的屋脊。不同海拔高度上温度、湿度和光照等多种环境因子的不同,使其草地物种多样性也存在着明显的差异。本研究以藏北地区那曲县念青唐古拉山脉日囤布山的高寒草地为试验样地,通过研究植物群落物种多样性的海拔梯度变化格局,分析藏北高寒草地植物群落特征及其物种多样性对海拔变化的响应,为该地区高寒草地生物多样性的保护和生态恢复提供科学依据和理论基础。
1 材料与方法
1.1 研究区概况 藏北(那曲)地区位于西藏冈底斯山和念青唐古拉山以北的广阔地区,83°41′~95°10′E,30°27′~35°39′N,是我国长江、怒江、澜沧江等主要江河的发源地之一[14]。土地总面积约为44.6万km2,占西藏自治区总面积的37.1%。该地区草地总面积约为42.1万km2,占整个地区土地总面积的94.4%[15]。丰富的草地资源和独特的气候条件、地理位置等,不仅对该区畜牧业发展以及生态系统的稳定性起着举足轻重的作用,而且对全国的江河、气候与环境都有直接或间接的影响,甚至对全球也有不容忽视的作用[16]。那曲县位于西藏自治区北部的青藏高原腹地,地处唐古拉山脉与念青唐古拉山脉之间。土地总面积160.44万hm2,总草地面积为138.72万hm2,占总面积的86.5%,是重要的畜牧业生产基地。该地区气候寒冷干燥,多大风,属亚寒带季风气候,平均气温-1.9℃,年降水量300~400mm,蒸发量1 799.6mm[17]。本试验样地选择在藏北地区那曲县念青唐古拉山脉日囤布山的高寒草地,其建群种主要为青藏苔草(Carex moorcroftii)、矮生嵩草(Kobresia humilis)、早熟禾(Poa annua)、紫花针茅(Stipapurpurea)、大花嵩草(K.macrantha)和鹅绒委陵菜(Potentilla anserina)等。
1.2 试验方法 于2009年8月,在草地植被生物量高峰期采用梯度格局,对那曲县念青唐古拉山脉日囤布山的高寒草地地上生物量及其植物群落物种多样性沿海拔梯度变化格局进行调查分析。由山脚(海拔高度为4 523m)至山顶(海拔高度为4 673 m)沿一条样线,每隔50m设置一个海拔梯度,共设置4个梯度,其分别为4 523、4 573、4 623、4 673m(表1),同时用GPS测量记录样地的经纬度。每一海拔梯度在约为50m×50m的样地上随机选取6个面积为0.5m×0.5m的样方,先记录每个样方内各个物种的高度、盖度和密度,然后齐地面分种收集地上生物量,称鲜质量后再置于105℃烘箱中杀青0.5h,以70℃恒温烘至质量不变,称干质量。
1.3 数据处理
1.3.1 各物种重要值[18]
物种重要值(IV)= (相对盖度+相对频度+相对高度)/3;
相对重要值(Pi)=IV/∑IV。

表1 样地基本信息
1.3.2 物种多样性 本研究中,物种丰富度用0.25 m2样方内出现的物种数表示。物种多样性的测定采用物种丰富度指数、Shannon-Wiener多样性指数(H)和E.Pielou均匀度指数(E)表征[19-21]。
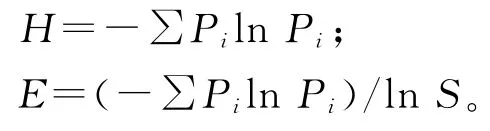
式中,S为群落物种数目,Pi为物种的相对重要值。1.4数据分析 用Excel和Access对数据进行初步整理,将各海拔梯度上所采的样方数据计算均值,通过SAS 9.0软件,利用单因子方差分析(ANOVA)和最小显著差数法(LSD)对不同海拔梯度下各指标间进行比较和差异显著性检验。
2 结果

图1 不同海拔梯度高寒草地植物群落的地上生物量及其盖度变化
2.1 不同海拔梯度上藏北高寒草地植物群落地上生物量及其盖度变化特征 随着海拔的逐渐升高,草地植被群落总盖度呈现先增加后降低的趋势(图1)。不同海拔梯度中,以海拔4 623m的植被盖度最高,超过了90%,与其他处理差异显著(P<0.05)。不同海拔梯度上,高寒草地植被地上生物量与总盖度的变化规律相似,即先增加后逐渐减少。随着海拔的升高,地上生物量鲜质量依次为47.73、48.00、90.40和33.07g/m2。其中在4 523与4 573m下几乎没有变化,而随着海拔继续升高,呈现先增加后降低的趋势。海拔4 623m与前两个梯度相比,鲜质量与干质量分别增加了88.3%~173%、33.8%~90.2%。由此可见,在藏北高海拔地区的气候环境中,不同海拔梯度对植物群落的盖度及其地上生物量影响显著,而海拔高度为4 623 m时,高寒草地植物群落的盖度和地上生物量均为最高,说明在该地区,中等海拔梯度(4 623m)水平最适宜草地植被的生长。
2.2 不同海拔梯度上高寒草地植物物种组成及其重要值的变化特征 通过对不同海拔梯度上植物物种重要值的分析,发现高寒草地植物群落物种组成及重要值存在很大的差异(表2)。在海拔4 523m处,建群种主要为青藏苔草和早熟禾等禾草类,伴生种有短穗兔耳草、海乳草以及少量的高原毛茛。由于该样地离居民点最近,草地出现了不同程度的退化,其植被组成较单一。随着海拔高度的增加,在海拔高度4 573m处,主要建群种为大花嵩草和矮生嵩草,次优势种为青藏苔草和早熟禾,伴生种主要为鹅绒委陵菜、肉果草和高山紫菀。在该海拔梯度,莎草类植物优势度明显增加,其中出现了矮生嵩草和大花嵩草等莎草科植物。在海拔高度4 623m处,草地植物物种种类最多,主要建群种和伴生种均发生了改变。其主要建群种为矮生嵩草、紫花针茅和鹅绒委陵菜,伴生种主要为矮小蓝钟花、滇紫草和星状风毛菊等杂类草。莎草类和杂类草的优势度总和在该梯度的比例明显增高。在4 673m海拔梯度下,建群种主要为矮生嵩草、鹅绒委陵菜和青藏苔草等,体现了随海拔高度的增加,杂类草的重要值表现出明显升高的趋势。
2.3 藏北高寒草地植物群落物种多样性沿海拔梯度的变化特征 不同海拔梯度上高寒草地植物群落物种丰富度、多样性和均匀度均具有显著差异(图2)。在海拔相对较低的层次物种丰富度较低;随着海拔高度的增加,物种丰富度呈先增加后减少的趋势。即海拔从4 523m升高至4 573m时,植物物种丰富度增加,至海拔4 623m处达到最高(15种);之后随海拔高度的继续增加,物种丰富度逐渐降低。多样性指数在4 623m海拔梯度下最高,为3.544;其次为海拔4 673m,其多样性指数为2.061;相对较低的海拔4 523与4 573m 处的多样性指数较低,分别为0.974和1.582。E.Pielou均匀度指数变化趋势与上述规律一致,即呈现海拔4 623m(1.309)>海拔4 673m(0.895)>海拔4 573m(0.813)>海拔4 523m(0.605)的变化格局。海拔高度与物种丰富度、多样性指数和均匀度指数之间的趋势模拟均呈负二次函数关系(图2),其 拟 合 系 数 r 值 分 别 为 0.825 8、0.825 2 和0.852 1。由分析可知,总体物种多样性随海拔高度的变化呈现“单峰”分布格局,即随着海拔的升高,物种丰富度、多样性和均匀度均先增加后降低,中等海拔高度(4 623m)时植物群落物种多样性最高。

表2 不同海拔梯度上高寒草地植物群落物种组成及其重要值的变化

图2 藏北高寒草地植物群落物种多样性沿海拔梯度变化趋势
3 讨论与结论
青藏高原独特的地理环境和气候条件,使该区域成为一个具有全球意义的脆弱生态系统[22]。海拔高度的变化首先引起温度、降水、大气成分等差异,从而影响土壤水分和太阳辐射照度等,进而影响植物群落组成和物种分布[23-24],因此,研究物种多样性的海拔梯度格局对于揭示生物多样性的环境梯度变化具有重要的意义。本研究表明,不同海拔梯度对草地植物群落生物量及其盖度具有显著的影响。青海玛沁县高寒草甸群落的地上生物量随着海拔的升高逐渐减少[25]。在本研究中,随着海拔的升高,高寒草地植物群落的总盖度、地上鲜生物量均表现出先增加后降低的趋势。原因可能是在海拔4 523 m高度下的草地离居民点较近,受人和牲畜的影响较大,植被稀疏,组成单一,且产量很低,该处的高寒草地已出现了严重的退化。随着海拔的升高,人为因素的干扰逐渐减少,草地群落优势种植物增多,生物量也明显增加,至4 623m海拔高度下达到最大值。随着海拔的进一步增高,环境因素逐渐成为草地生长的主要限制因子,群落生物量及其盖度显著下降。综合表明,在本研究中海拔高度4 623m水平最适应草地植物的生长。高寒草地植物群落物种组成及其重要值沿海拔梯度变化存在很大的差异。随着海拔的升高,植被群落中出现了莎草类和杂草类植物,而且其重要值所占的比例也明显增加。
由于影响草本植物分布的因素较多,草地植物群落物种多样性随海拔梯度变化的规律非常复杂,不同研究尺度上得出的结果不尽相同[4,26]。贺金生和陈伟烈[26]将物种多样性随海拔梯度的变化归纳为5种形式,分别为:随着海拔的升高而降低(负相关)、先升高后降低(中间高度膨胀型)、先降低后升高、单调升高(正相关)和无规律性(无关)。在本研究中,高寒草地总体物种多样性随海拔变化呈现“单峰”分布格局,即随着海拔的升高,物种丰富度、Shannon-Wiener多样性指数和E.Pielous均匀度指数均表现出先增加后降低的趋势,中海拔高度植物群落物种多样性最高,即“中间高度膨胀型”,这与王国宏[27]在祁连山区的研究结论相一致。在低海拔区域,较频繁的人为干扰、破坏和降水的限制导致该区域物种多样性较低[28]。一般而言,随着海拔高度的增加,气温直减率为每100m降低0.5~0.7℃[23]。在高海拔地区,气温的逐渐降低使非耐寒草本物种受到热量的限制而减少,物种多样性也相应降低。而在中等海拔高度,一方面,由于没有人为干扰和破坏,草地植物群落生长旺盛;另一方面,由于具有植物所需要的适宜的水热配置,该区域具有丰富的物种多样性分布[10,29]。本研究中那曲县唐古拉山脉日囤布山低海拔区域由于接近居名点,一定程度上受到人为因素的干扰,物种多样性相对较低,因此,低海拔区域草地植被的保护对于草地生产力的维持和生物多样性的稳定发展具有重要的意义。
在全球气候变化背景下,研究物种多样性随海拔梯度的变化在一定程度上可以用于探讨物种多样性对极端气候变化的响应与适应。基于青藏高原气候的复杂性和脆弱性等特征,开展高寒草地恢复与生物多样性保护等方面的研究面临着诸多的问题和挑战。由于受到青藏高原这一特殊生境的影响,物种多样性沿海拔梯度变化格局可能还受到其他环境因子的影响。本研究可能还受到研究区相对海拔高度的限制,由于没有足够长的梯度,分布格局变化可能具有不确定的因素[30]。因此,开展不同尺度的生物多样性的研究以及它们对全球气候变化的响应,是未来研究的重要方向。
[1] 周兴民.生物多样性的研究与保护[J].青海环境,1996(4):19-21.
[2] Tilman D,Wedin D,Knops J.Productivity and sustainability influenced by biodiversity in grassland ecosystems[J].Nature,1996,379:718-720.
[3] Tilman D,Reich P B,Knops J,et al.Diversity and productivity in a long-term grassland experiment[J].Science,2001,294:843-845.
[4] 唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20-28.
[5] 杜岩功,崔骁勇,葛劲松,等.三江源地区高寒草地群落特征研究[J].草业科学,2010,27(3):9-14.
[6] Gaston K J.Global patterns in biodiversity[J].Nature,2000,405:220-226.
[7] Oommen M A,Shanker K.Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants[J].Ecology,2005,86:3039-3047.
[8] 郝占庆,于德永,杨晓明,等.长白山北坡植物群落α多样性及其随海拔梯度的变化[J].应用生态学报,2002,13(7):785-789.
[9] 唐志尧,方精云,张玲.秦岭太白山木本植物物种多样性的梯度格局及环境解释[J].生物多样性,2004,12(1):115-122.
[10] Wang G H,Zhou G S,Yang L M,et al.Distribution,species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains,Gansu,China[J].Plant Ecology,2002,165:169-181.
[11] 王国宏.祁连山北坡中段植物群落多样性的垂直分布格局[J].生物多样性,2002,10(1):7-14.
[12] 杨元合,饶胜,胡会峰,等.青藏高寒草地植物物种丰富度及其与环境因子和生物量的关系[J].2004,12(1):200-205.
[13] 贺连选,刘宝汉.不同海拔高度的高寒草甸植物群落多样性的研究[J].青海畜牧兽医杂志,2005,35(5):1-4.
[14] 高清竹,万运帆,李玉娥,等.藏北高寒草地NPP变化趋势及其对人类活动的响应[J].生态学报,2007,27(11):4612-4619.
[15] 甘肃草原生态研究所,西藏那曲地区畜牧局.西藏那曲地区草地畜牧业资源[M].兰州:甘肃科学技术出版社,1991:14-33.
[16] 高清竹,李玉娥,林而达,等.藏北地区草地退化的时空分布特征[J].地理学报,2005,60(6):965-973.
[17] 西藏自治区土地管理局,西藏自治区畜牧局.西藏自治区草地资源[M].北京:科学出版社,1994.
[18] 蒙旭辉,李向林,辛晓平,等.不同放牧强度下羊草草甸草原群落特征及多样性分析[J].草地学报,2009,17(2):239-244.
[19] Zhou Z,Sun O J,Huang J,et al.Land use affects the relationship between species diversity and productivi-ty at the local scale in a semi-arid steppe ecosystem[J].Functional Ecology,2006,20:753-762.
[20] Pielou E C.Mathematical Ecology[M].New York:John Wiley & Sons.Interscience,1977:291-311.
[21] 袁建立,江小蕾,黄文冰,等.放牧季节及放牧强度对高寒草地植物多样性的影响[J].草业学报,2004,13(3):16-21.
[22] 曹广民,林丽,张法伟,等.青藏高原高寒矮嵩草草甸稳定性的维持、丧失与恢复[J].草业科学,2010,27(8):34-38.
[23] Fonda R W,Bliss L C.Forest vegetation of the montane and subalpine zones,Olympic Mountains,Washington[J].Ecological Monographs,1969,39:271-301.
[24] 方精云,沈泽昊,崔海亭.试论山地的生态特征及山地生态学的研究内容[J].生物多样性,2004,12(1):10-19.
[25] 王长庭,王启基,龙瑞军,等.高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究[J].植物生态学报,2004,28(2):240-245.
[26] 贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17(1):91-99.
[27] 王国宏.祁连山北坡中段植物群落多样性的垂直分布格局[J].生物多样性,2002,10(1):7-14.
[28] 冯建孟,王襄平,徐成东,等.玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局[J].山地学报,2006,24(1):110-116.
[29] 朱源,康慕谊,江源,等.贺兰山木本植物群落物种多样性的海拔格局[J].植物生态学报,2008,32(3):574-581.
[30] Whittaker R H,Willis K J,Field R.Scale and species richness:towards a general,hierarchical theory of species diversity[J].Journal of Biogeography,2001,28:453-470.
