盐胁迫对两种锦鸡儿幼苗生理指标的影响
2011-08-20苗昊翠李利民黄俊华
苗昊翠,李利民,宋 彬,黄俊华
(1.新疆农业科学院农作物品种资源研究所,新疆 乌鲁木齐830091;2.新疆农业大学林学与园艺学院,新疆 乌鲁木齐830052)
目前,全世界有100多个国家3.8亿hm2土地受盐碱化影响,约占陆地面积的1/3,土壤盐碱化已成为世界农业生产面临的主要威胁之一[1]。新疆盐碱地具有面积大、类型多、积盐重、形成复杂等特点,各类盐渍化土地面积约11万km2,北疆以硫酸盐或硫酸盐+氯化物为主,南疆以氯化物或氯化物+硫酸盐为主[2]。盐分是影响植物生长和产量的一个重要环境因子,盐胁迫会使植物生长量降低甚至死亡[3]。植物对盐分胁迫的反应和适应是一个复杂的生理过程,是植物体内一系列生理生化过程综合作用的结果[4-6]。新疆盐渍荒漠蒸发强、降水少、地表积盐经常存在且含盐量随季节变化,植物生长的整个时期都处于盐分状态下,因此,植物种子的萌发和幼苗生长对盐生环境的适应能力是决定盐生植物种群更新的关键因素[7]。近年来,国内外对植物和盐胁迫的关系进行了大量的研究[8-10],但有关锦鸡儿属植物耐盐性研究较少。
锦鸡儿属植物隶属于豆科,该属模式种为树锦鸡儿(Caragana arborescens)。全世界有100余种,大致分布在28°~56°N[少数种类如鬼箭锦鸡儿(C.jubata)分布至73°N],30°~140°E的地带。我国产62种,9变种,12变型[11],新疆有32种,种类约占全国种类的1/2,占全世界的1/3,是中国锦鸡儿属种类多样性最多的地区,也是世界锦鸡儿属种类最集中的地区之一,具有明显的种类优势。锦鸡儿属植物抗旱性强,在北方干旱地区防风固沙、保持水土、改善土壤养分中,发挥着良好的生态效益。
对锦鸡儿属植物的研究现多集中于分类、分布、栽培及抗旱性等方面,对抗盐特性的研究很少,本研究经过调查与分析,选择了广泛分布于新疆荒漠的白皮锦鸡儿(C.leucophloea)和从山前平原到戈壁滩皆有分布的镰叶锦鸡儿(C.aurantiaca)为研究对象。对两种锦鸡儿盐分胁迫下幼苗期生理指标变化的研究,不仅有利于锦鸡儿属植物抗性资源的开发利用,而且对新疆大面积盐渍土壤的改良、开发和利用,改善日益脆弱的生态环境,以及西北城市绿化建设有着重要的意义。
1 材料与方法
1.1 试验地及供试材料 本试验在新疆乌鲁木齐县种苗场试验地进行。试验材料为白皮锦鸡儿、镰叶锦鸡儿,种子分别采自新疆柴窝堡林场(43°33.285′N,87°52.108′E,海拔1 124m)、新疆伊犁昭苏军马场河谷(43°07.316′N,80°59.870′E,海拔1 881m)的自然植物群落中,分别从各坡向的株丛上采种并混和。
1.2 研究方法
1.2.1 盆栽试验设计 试验设为单因素即不同的NaCl质量分数的处理,设0.2%、0.4%、0.6%、0.8%和1.0%共5个盐质量分数梯度,对照为正常生长;在胁迫后7和15d进行取样,测定其生理指标。每个处理设5个重复,每种植物材料为60盆,共120盆。用塑料花盆装土,每盆装土1.5kg。试验选取生长3个月,平均株高20cm的健康幼苗将预先称好的NaCl用适量自来水溶解,按照不同盐质量分数对各个植物材料依次施入盆中,对照加等量的自来水,以后每天称量,及时补充蒸散掉的水分,尽量保持盐质量分数的恒定,减少试验误差。
1.2.2 膜透性测定 采用电导法测定[12],以相对电导率表示。将新鲜锦鸡儿叶样用蒸馏水冲洗3次,用滤纸吸干表面水分,分别称取0.3g放入小烧杯中,加入蒸馏水15mL,静置30min,用EC214型电导仪测其电导值(L1),然后置于沸水浴中5min以杀死组织,待冷至室温后,测电导值(L2)。
相对电导率=L1/L2×100%。
1.2.3 丙二醛(MDA)和游离脯氨酸含量的测定丙二醛含量采用硫代巴比妥酸(TBA)方法测定[13]。游离脯氨酸含量采用酸性茚三酮法测定[13]。
1.2.4 统计分析 所得数据用SPSS 12.0软件进行统计分析,Excel软件绘图。
2 结果与分析
2.1 盐胁迫对两种锦鸡儿幼苗茎叶细胞膜透性的影响 在不同盐胁迫条件下,白皮锦鸡儿的相对电导率均低于镰叶锦鸡儿(图1)。膜透性的大小,反映质膜受伤害的程度;数值越大,质膜受到的伤害也越大,说明白皮锦鸡儿质膜受伤害程度低于镰叶锦鸡儿。
不同质量分数NaCl胁迫7d,两种锦鸡儿叶片膜透性均发生了一定的变化(图1)。随NaCl质量分数的增加,镰叶锦鸡儿叶片相对电导率有所增加,0.2%NaCl时达到最大,后降低并趋于稳定;白皮锦鸡儿叶片相对电导率较镰叶锦鸡儿低,随NaCl质量分数的增加,白皮锦鸡儿的相对电导率与对照无显著差异(P>0.05),基本表现出稳定的趋势,说明白皮锦鸡儿叶在NaCl胁迫7d时没有受到明显伤害。
不同质量分数NaCl胁迫15d,白皮锦鸡儿叶片在不同盐质量分数胁迫下叶片相对电导率与对照差异不显著(P>0.05),镰叶锦鸡儿叶片相对电导率总体随盐质量分数的增大逐渐升高。与7d相比,白皮锦鸡儿叶片相对电导率表现出各NaCl胁迫质量分数下15d低于7d,这可能是因为白皮锦鸡儿植株生理活动旺盛,加之大气温度和湿度条件适宜,而使其幼苗对盐胁迫适应能力增强;镰叶锦鸡儿叶片相对电导率表现为15d高于7d,但增幅不明显。
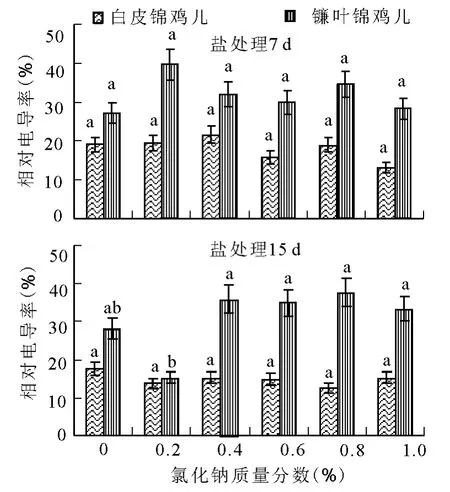
图1 盐胁迫对两种锦鸡儿叶片相对电导率的影响
2.2 盐胁迫对两种锦鸡儿幼苗丙二醛含量的影响 受不同质量分数NaCl胁迫的影响,两种锦鸡儿幼苗叶片的MDA含量均发生了一定的变化。胁迫7d的幼苗,白皮锦鸡儿表现出MDA的含量低于对照组并基本保持稳定(图2),在0.2%NaCl中幼苗的MDA含量最低,后随NaCl质量分数的升高,MDA的含量缓慢升高并趋于稳定;镰叶锦鸡儿MDA随NaCl质量分数的升高呈先降低后升高的趋势,在NaCl质量分数1.0%胁迫下达到最高。
NaCl胁迫15d的幼苗,随NaCl质量分数的增加,白皮锦鸡儿MDA含量呈升高-降低-升高的趋势,在NaCl为0.2%时达到最大值,不同盐质量分数胁迫与对照均有显著差异(P<0.05);镰叶锦鸡儿MDA含量在NaCl为0.2%~0.6%时与对照差异不显著(P>0.05),当 NaCl为0.8%~1.0%时与对照有显著差异(P<0.05)(图2)。
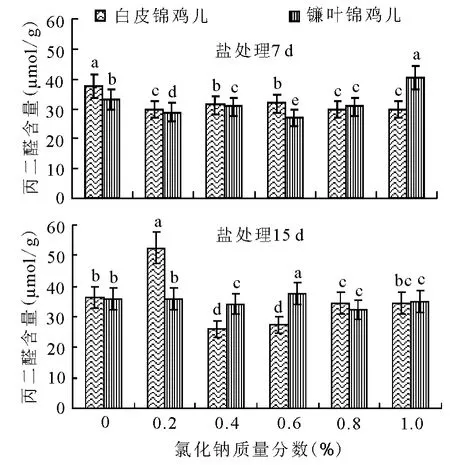
图2 盐胁迫对两种锦鸡儿叶丙二醛的影响
NaC1胁迫15d与7d相比,白皮锦鸡儿总体表现出随胁迫时间的延长MDA含量在NaCl质量分数为0.4%、0.6%时有所降低,在NaCl质量分数为0.8%、1.0%时有所增加;镰叶锦鸡儿表现为胁迫15dMDA含量较7d高。
2.3 盐胁迫对两种锦鸡儿幼苗游离脯氨酸含量的影响 白皮锦鸡儿脯氨酸含量除在1.0%盐胁迫7d时显著低于镰叶锦鸡儿(图3),其他胁迫条件下均高于镰叶锦鸡儿或基本相同。

图3 盐胁迫对两种锦鸡儿叶游离脯氨酸含量的影响
盐胁迫7d时,白皮锦鸡儿叶片中脯氨酸含量除在NaCl为0.6%时有所升高外,其余胁迫条件下的脯含酸含量均低于对照。而镰叶锦鸡儿叶片脯氨酸含量在NaCl质量分数低于0.6%时变化不大,在NaCl质量分数达到0.8%时,脯氨酸含量出现明显的升高,尤其在1.0%质量分数下,脯氨酸含量急剧增加,这表明镰叶锦鸡儿叶片在较高质量分数盐胁迫下需要通过积累脯氨酸来提高细胞的渗透调节能力,从而保证正常的细胞膨压[14]。
脯氨酸的积累是对盐逆境适应性的表现[2],15 d时两种锦鸡儿脯氨酸含量较7d时均有所增加,这说明随着胁迫时间的延长和盐胁迫程度的增加,两种锦鸡儿通过增加体内脯氨酸含量维持渗透平衡,适应盐逆境。盐胁迫15d,在低盐0.2%胁迫处理时白皮锦鸡儿与镰叶锦鸡儿脯氨酸含量均显著增加(P<0.05),随盐质量分数提高,白皮锦鸡儿叶片脯氨酸含量趋于稳定,而镰叶锦鸡儿在0.4%盐溶液胁迫下再次出现显著升高,后逐渐趋于平稳,说明随胁迫时间的延长,在低质量分数盐溶液下两种锦鸡儿均开始产生脯氨酸适应盐胁迫。但与对照相比,白皮锦鸡儿叶片脯氨酸含量在各盐溶液质量分数梯度下增加幅度均明显低于镰叶锦鸡儿。
3 讨论
3.1 盐胁迫对两种锦鸡儿幼苗膜透性的影响
细胞膜是外界盐离子进入细胞的第一道屏障[15],在逆境条件下,植物叶片的膜透性能反映植物细胞膜系统的损伤程度[16-17]。高盐质量分数能增加细胞膜的透性,加强膜质过氧化作用,最后导致膜系统破坏[18-19]。因此细胞膜透性研究是植物抗逆性研究的重要部分之一。当质膜的选择透性因逆境伤害而明显改变或丧失时,细胞内的物质大量外渗,导致组织浸泡液的电导率增大,通过测定外渗液电导率的变化,就可以反映出质膜受伤害的程度和所测材料抗逆性的大小。本研究表明,随着NaCl胁迫程度的增加及胁迫时间的延长,白皮锦鸡儿叶细胞质膜的相对透性表现出逐渐降低,而镰叶锦鸡儿的膜透性有所增加,说明白皮锦鸡儿耐盐性较镰叶锦鸡儿强。
3.2 盐胁迫对两种锦鸡儿幼苗丙二醛的影响
丙二醛是膜脂过氧化作用的产物之一[20],它可与膜蛋白发生交联作用,使膜透性增大;又可与细胞内的各种成分发生反应,使膜系统中多种酶的生理功能严重受损。因此,可用丙二醛含量来代表植物膜脂过氧化水平,反映植物受伤害的程度[21]和植物对逆境的反应[22]。植物在逆境下往往发生膜脂过氧化作用,破坏细胞膜的结构,积累许多有害的过氧化物,丙二醛作为膜脂过氧化水平的指标已为人们所接受[23]。本研究显示,白皮锦鸡儿随盐胁迫时间的延长表现出在低质量分数盐(0.4%~0.6%)胁迫下丙二醛含量有所下降,在高质量分数盐(0.8%~1.0%)胁迫下丙二醛含量有所升高,这一现象说明白皮锦鸡儿在低盐质量分数胁迫下,通过自身调节机制逐步适应新的环境,而高盐质量分数损伤了膜系统;还表现出NaCl为0.4%~1.0%时丙二醛含量逐渐升高,但均低于对照,这种现象与尹增芳等[14]在海滨锦葵(Kosteletzkya virginica)幼苗中的发现相似,说明低盐溶液可降低丙二醛在植物体内的积累,而NaCl质量分数的升高可诱导丙二醛含量的增加。镰叶锦鸡儿表现出随盐胁迫时间的延长丙二醛含量升高,说明植物体内受到一定的伤害,引起膜的过氧化,导致丙二醛含量升高。有研究[15]表明,不同棉花(Gossypium hirsutum)耐盐品种丙二醛积累量反映出其耐盐性的差异,耐盐性较低的品种其植株体内的丙二醛含量较高。该指标结果说明白皮锦鸡儿耐盐性高于镰叶锦鸡儿。
3.3 盐胁迫对两种锦鸡儿幼苗脯氨酸的影响
脯氨酸是植物体内有机渗透物质,其作用可以概括为:调节细胞质的渗透势;保护蛋白质分子,增加蛋白质分子的水合度;维持光合活性;作为活性氧的清除剂等[23]。对于某些植物而言,几乎所有的逆境,包括干旱、低温、高温、冰冻、盐害都可以造成体内脯氨酸的累积[24-25],脯氨酸的积累是植物为了对抗盐胁迫而采取的一种保护性措施[26-27]。
从本研究结果看,盐胁迫7d对白皮锦鸡儿的影响不大,而镰叶锦鸡儿在较高盐溶液下出现明显的脯氨酸积累。而随着盐胁迫时间的延长,两种锦鸡儿虽然都在低盐溶液下表现出叶片脯氨酸含量增加,但镰叶锦鸡儿叶片脯氨酸含量增幅远超过白皮锦鸡儿。说明在这一指标中,镰叶锦鸡儿对盐胁迫表现的更加敏感。
肖雯等[28]通过对11种盐生植物与3种非盐生植物抗盐生理指标的研究发现,盐生植物组的功能叶中膜透性平均值低于对照组,脯氨酸含量在所测3种渗透调节剂中所占比例最高,而且盐生植物组平均值高于对照组;孙黎等[29]通过对10种藜科盐生植物与2种非盐生植物的抗盐生理生化特征的研究也得出相同的结论。本研究中不同胁迫处理的白皮锦鸡儿叶片脯氨酸含量多高于镰叶锦鸡儿或基本相同,说明白皮锦鸡儿较镰叶锦鸡儿更耐盐。脯氨酸含量的变化与盐胁迫程度成相关性,随着盐胁迫质量分数的增大,植物体内的脯氨酸含量也逐渐增加,表明它可能在帮助植物抵御逆境以及增强耐盐性方面起到重要作用。
综合比较两种锦鸡儿叶片相对电导率、丙二醛和脯氨酸含量可知,随着盐质量分数的增加,胁迫时间的延长,两种锦鸡儿总体上在各项生理指标中均表现出一定的生理响应,白皮锦鸡儿表现出较镰叶锦鸡儿更耐盐,3项指标中细胞膜透性与脯氨酸含量对于两种锦鸡儿的抗盐性具有明显的指示意义。
[1] 夏尚光,张金池,梁淑英.NaCl胁迫对3种榆树幼苗生理特性的影响[J].河北农业大学学报,2008(2):52-56.
[2] 克热木·伊力,袁琳,齐曼·尤努斯,等.盐胁迫对阿月浑子SOD、CAT、POD活性的影响[J].新疆农业科学,2004,41(3):129-132.
[3] 裘丽珍,黄友军,黄坚钦,等.不同耐盐性植物在盐胁迫下的生长与生理特性比较研究[J].浙江农业大学学报(农业与生命科学版),2006,32(4):420-427.
[4] 杨少辉,季静,王罡,等.盐胁迫对植物影响的研究进展[J].分子植物育种,2006,4(3):139-142.
[5] Cramer G R,Lauchli A,Epstein E.Effects of NaCl and CaCl2on ion activities in comple nutrient and root growth of cotton[J].Plant Physiololy,1986,81:792-797.
[6] 王素平,郭世荣,胡晓辉,等.盐胁迫对黄瓜幼苗根系生长和水分利用的影响[J].应用生态学报,2006,17(10):1883-1888.
[7] 渠晓霞,黄振英.盐生植物种子萌发对环境的适应策略[J].生态学报,2005,25(9):2389-2398.
[8] Santa-Cruz A,Acosta M,Rus A,et al.Short-term salt tolerance mechanisms in differentially salt tolerant tomato species[J].Plant Physiology and Biochemistry,1999,37(1):65-71.
[9] Saneoka H,Nagasaka C,Hahn D T,et al.Salt tolerance of glycinebetaine-deficient and containing maize lines[J].Plant Physiology,1995,107:631-638.
[10] Soussi M,Ocana A,Lluch C.Effects of salt stress on growth,photosynthesis and nitrogen fixation in chickpea(Cicer arietinumL.)[J].Journal of Experimental Botany,1998,49(325):1329-1337.
[11] 中国科学院中国植物志编辑委员会.中国植物志(第四十二卷,第一分册)[M].北京:科学出版社,1993.
[12] 邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000.
[13] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[14] 尹增芳,何祯祥,王丽霞,等.NaCl胁迫下海滨锦葵种子萌发和幼苗生长过程的生理特性变化[J].植物资源与环境学报,2006,15(1):14-17.
[15] 孙小芳,郑青松,刘友良.NaCl胁迫对棉花种子萌发和幼苗生长的伤害[J].植物资源与环境学报,2000,9(3):22-25.
[16] 蒋明义,荆家海,王韶唐.渗透胁迫对水稻幼苗膜脂过氧化及体内保护系统的影响[J].植物生理学报,1991,17(1):80-84.
[17] 陈立松,刘星辉.水分胁迫对龙眼幼苗叶片膜脂过氧化及内源保护体系的影响[J].武汉植物学研究,1999,17(2):105-109.
[18] 郑丽锦,张学英.果树盐胁迫生理生化特性的研究进展[J].河北农业大学学报,2003,26(3):41-44.
[19] 辛华,张秀芬,初庆刚.山东滨海盐生植物叶结构的比较研究[J].西北植物学报,1998,18(4):584-589.
[20] 周学丽,周青平,颜红波,等.NaCl胁迫对同德小花碱茅苗期生理特性的影响[J].草业科学,2009,26(6):101-105.
[21] 周革,倪福太,张立娟.植物在逆境中的形态结构及生理变化[J].吉林师范大学学报,2004(2):64-67.
[22] 袁琳,克热木.NaCl胁迫对阿月浑子实生苗活性氧代谢与细胞膜稳定性的影响[J].植物生态学报,2005,29(6):985-991.
[23] 冯大千.旱生观赏植物资源及其开发利用价值[J].八一农学院学报,1991,14(3):76-82.
[24] 朱小龙,李振基,赖志华,等.不同光照下土壤水分胁迫对长苞铁杉幼苗的作用[J].北京林业大学学报,2007,29(3):76-81.
[25] 李源,刘贵波,高洪文,等.紫花苜蓿种质耐盐性综合评价及盐胁迫下的生理反应[J].草业学报,2010,19(4):79-86.
[26] 陈托兄,张金林,陆妮,等.不同类型抗盐植物整株水平游离脯氨酸的分配[J].草业学报,2006,15(1):36-41.
[27] 公勤,齐曼·尤努斯,艾力江·买买提.NaCl胁迫对3种胡颓子属植物幼苗体内物质积累及水分含量的影响[J].新疆农业大学学报,2008,31(3):46-50.
[28] 肖雯,贾恢先,蒲陆梅.几种盐生植物抗盐生理指标的研究[J].西北植物学报,2000,20(5):818-825.
[29] 孙黎,刘士辉,师向东,等.10种藜科盐生植物的抗盐生理 生 化 特 征 [J].干 旱 区 研 究,2006,23(2):309-313.
