中国南方不同品系福寿螺对广州管圆线虫易感性的研究*
2011-06-07曾清香孙志坚彭东觉曲振宇吴忠道詹希美
曾清香,孙志坚,张 波,彭东觉,曲振宇,杨 萧,吴忠道,詹希美,何 蔼
福寿螺(Pomacea canaliculata,Lamarck 1822)原产于南美洲,属外来入侵物种[1],其可作为广州管圆线虫(Angiostrongylus cantonensis,Chen 1935)的中间宿主引起人嗜酸性脑膜脑炎[2]。自从Nomura和 Lin[3]报道了首例人感染广州管圆线虫以来,至1992年已经有大约30个国家有病例的报道[4]。而在中国,在1996年之前,广州管圆线虫感染的病例只有3例,而近些年来,浙江、福建、北京等地先后出现广州管圆线虫病暴发,使得此数目迅速增加[5]。在几次广州管圆线虫大暴发流行中,均与福寿螺有关,Prociv等[6]甚至认为福寿螺已经成了印度洋-太平洋区广州管圆线虫病流行的首要因素。
有关福寿螺壳的变异,早在19世纪就有人注意到,由于福寿螺适应能力强,分布范围广,其摄食、繁殖、活动的情况都可能在不同地区有不同的变化[7],因此其变异应该会更多[8]。2000年中国就有人指出福寿螺可分为黑色福寿螺和黄色福寿螺[9],我们在对中国广州各地福寿螺的观察中发现,除了上述依据壳色分类的两类福寿螺外,还存在另外一类。依照传统的命名,我们将其称为灰色福寿螺,这是通过对福寿螺外壳和软体部综合观察得出的[10]。本实验主要研究了三类福寿螺对广州管圆线虫的易感性。
1 材料与方法
三类福寿螺的外形特点及来源[10]外形特点:(1)黑色福寿螺(见图1a):壳为棕绿色,上有数量不等的黑色色带,厣为深褐色,软体部包括唇须、触角、呼吸管、足等均为黑色;(2)黄色福寿螺(见图1b):壳为棕黄色,不含色带,厣亦为金黄色,软体部包括唇须、触角、呼吸管、足等均为黄白色;(3)灰色福寿螺(见图1c):壳的外观肉眼见与黑色福寿螺无区别,但软体部,尤其在唇须、触角、呼吸管、足边缘可见大量黄色斑点。
依据以上外形特点采集福寿螺样本,大部分黑色福寿螺来自中国科学院华南植物园池塘,少量来自华南农业大学试验田水渠;大部分黄色福寿螺来自华南农业大学试验田水渠,少部分来自广州农业科学研究所和中国华南植物园;所有灰螺均来自广州农业科学研究所。

图1 3类福寿螺外部形态a黑色福寿螺、b黄色福寿螺、c灰色福寿螺。上图显示螺壳形态,下图显示爬行时外露软体部形态。Fig.1 External features of three types of P.canaliculata1a,1b and 1c represented black P.canaliculata,yellow P.canaliculata and grey P.canaliculata respectively.And the upper showed the shells and the lower part showed the soft parts when they was crawling
1.2 广州管圆线虫Ⅰ期幼虫(First stage larvae of A.cantonensis,L1)的获得 将野外感染广州管圆线虫的褐云玛瑙螺(采自广州大学城)去壳,软组织剁碎后置于人工消化液(按2g胃蛋白酶、7mL浓盐酸、1 000mL蒸馏水配制)内,于 37℃温箱中消化90min并在解剖镜下分离广州管圆线虫三期幼虫(Third stage larvae of A.cantonensis,L3),将检出的L3活虫40条灌胃感染SD大白鼠,6周后,收集新鲜鼠粪并用Baermann技术[11]分离L1。
1.3 福寿螺感染过程 预先将3类福寿螺各100只按上述消化褐云玛瑙螺的方法消化并查找L3,结果均无广州管圆线虫感染,证明实验用福寿螺均无野外感染。取三类福寿螺各90只,每类再随机分为3组,每组30只。分别记为黑色福寿螺Ⅰ组、Ⅱ组、Ⅲ组;黄色福寿螺Ⅰ组、Ⅱ组、Ⅲ组;灰色福寿螺Ⅰ组、Ⅱ组、Ⅲ组。各组螺于感染前两天停止喂食。Ⅰ组作为阴性对照组,不感染广州管圆线虫;Ⅱ、Ⅲ组分别为低剂量感染组和高剂量感染组,分别用每只螺500条和2000条L1的感染量感染福寿螺24 h,24 h后再使用相同剂量重复感染1次,即相当于低剂量感染组感染量为每只螺1 000条L1,高剂量组为4 000条。
感染时,将各组福寿螺置于相同规格的水盆中,加水至恰能淹没所有福寿螺,再加入预定数量的L1(对照组加入相同体积的水),使福寿螺在含L1的水中感染24h。感染完成后,将所有螺置于有流动水的大池中,喂以足量蔬菜。35d后,将所有螺取出,分别去壳取软组织,将软组织剁碎后每克螺肉加入20mL人工消化液(同上),置于37℃温箱中消化90min。在解剖显微镜下查找L3,按照Hata等[12]的描述对L3进行鉴定,并计数。
1.4 观察指标 实验前称取各组福寿螺重量,分别于感染前、感染后、感染后7、14、21、28和35 d观察各组福寿螺活动度,观察并记录整个过程中螺的死亡数量,称取实验后各组福寿螺重量,记录各螺感染L3数,并计算感染率、感染度、虫密度(感染度/螺重量)。感染度为每只福寿螺感染L3的数量。
福寿螺活动度的观察:将福寿螺取出置于水盆中,加水至淹没所有螺,摇晃水盆以使所有螺均退回壳内。观察并记录 1min、3min、5min和10min开始活动的福寿螺数量,并计算活动率(活动螺数/总螺数)。福寿螺活动以伸出呼吸管或露出触角或开始爬行为标准。
1.5 数据统计 使用SPSS13.0对数据进行统计。福寿螺实验前后重量用t检验;死亡率及感染率运用χ2检验;感染度及虫密度运用秩和检验;运用spearman相关分析,对感染L3福寿螺重量及感染度进行相关分析。
2 结 果
2.1 实验前后实验组福寿螺重量 分别称取了实验组实验前后各福寿螺的重量,见表1。运用T检验对各组福寿螺实验前后重量进行两两比较,均无差别(P>0.05)。说明在实验的30余天中,福寿螺并无生长。在同一组中,灰色福寿螺均小于黑色福寿螺和黄色福寿螺(P<0.05),在Ⅲ组中,黑色福寿螺小于黄色福寿螺(P<0.05),而在Ⅱ组中,黑色福寿螺与黄色福寿螺重量无差异(P>0.05)。
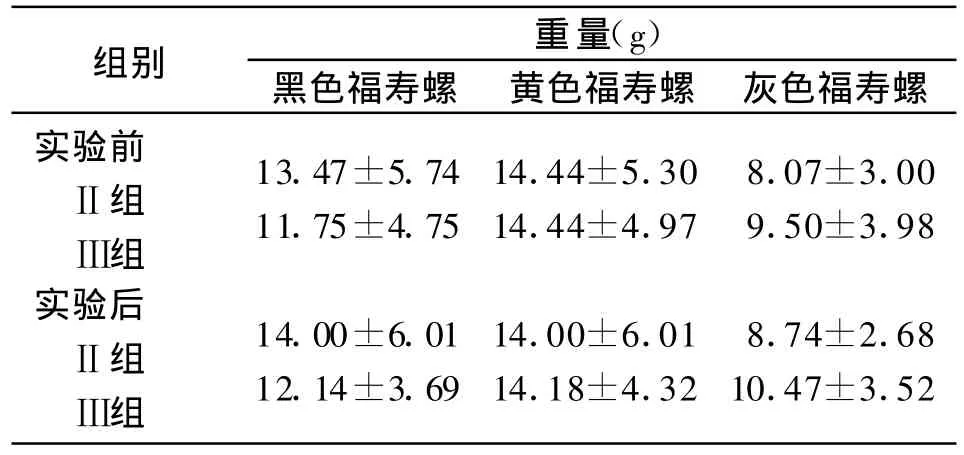
表1 实验前后实验组福寿螺重量Table 1 Comparison of snails'weights between pre-and postinfection
2.2 福寿螺活动度的变化 福寿螺活动率变化幅度较大,可能与外界环境如当日温度、天气等不同有关。但同阴性对照组相比,可见感染组活动率在感染完成后和感染后7 d明显下降,随后逐渐上升,尤以Ⅱ组明显,至感染后28 d时,实验组与Ⅰ组差别已不明显;另外,Ⅲ组活动率较Ⅱ组下降明显(见图2a)。相同组中,三类螺活动率无明显规律(见图2b)。
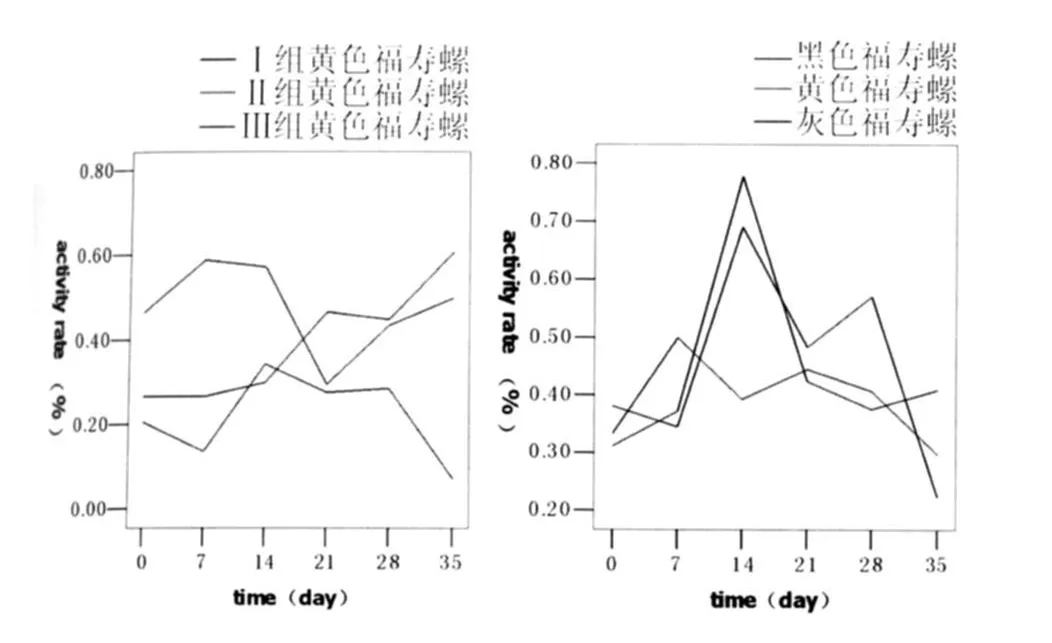
图2a 各组黄色福寿螺不同时间5min时活动率图2b Ⅲ组不同类福寿螺5min时活动率Fig.2a Activity rate of yellow P.canaliculata at 5thminuteFig.2b Activity rate of dif ferent P.canaliculata in GroupⅢat 5thminute
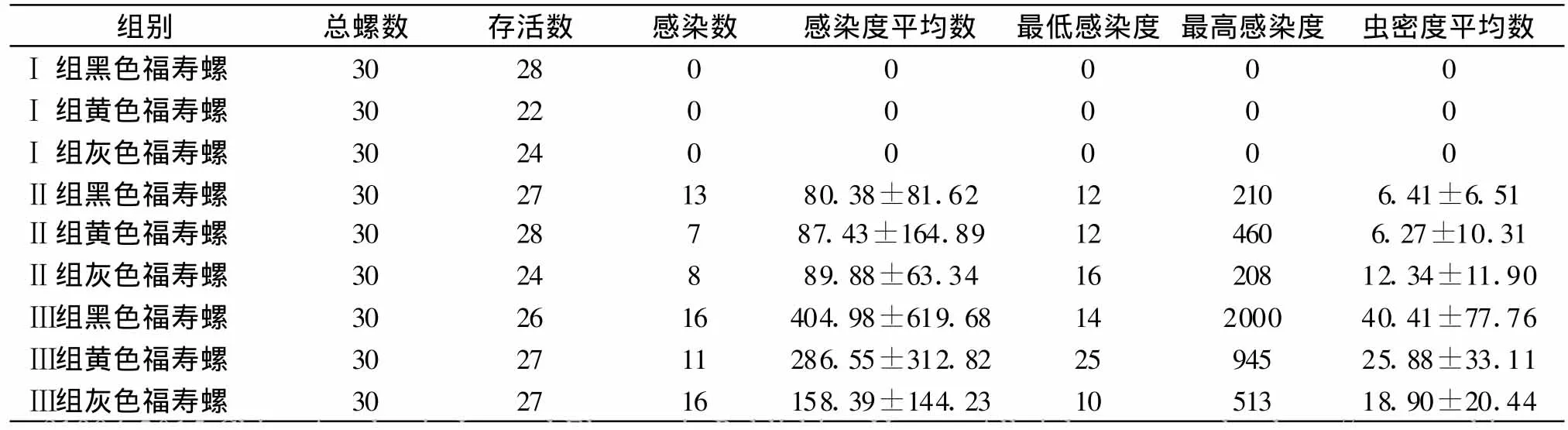
表2 各组福寿螺死亡及感染情况Table 2 Infection status of different groups
2.3 死亡率 实验结束时,黑色福寿螺死亡率Ⅰ组为 6.67%(2/30)、Ⅱ组为 10%(3/30)、Ⅲ组为13.3%(4/30);黄色福寿螺死亡率Ⅰ组为26.67%(8/30)、Ⅱ组为6.67%(2/30)、Ⅲ组为10%(3/30);灰色福寿螺死亡率Ⅰ组为20%(6/30)、Ⅱ组为20%(6/30)、Ⅲ组为30%(3/30),见表 2。运用 χ2检验,黑色福寿螺各组两两比较死亡率无差异(P>0.05);黄色福寿螺Ⅰ组死亡率高于Ⅱ组和Ⅲ组(P<0.05),Ⅱ组和Ⅲ组之间比较无统计学意义(P>0.05);灰色福寿螺各组两两比较死亡率无差别(P>0.05)。运用χ2检验对相同组不同类螺死亡率进行检验得出,Ⅰ组黄色福寿螺死亡率高于Ⅰ组黑色福寿螺(P<0.05),而与Ⅰ组灰色福寿螺比较无明显差异(P>0.05),Ⅰ组黑色福寿螺与灰色福寿螺比较无差别(P>0.05);Ⅱ组中三类福寿螺死亡率两两比较无统计学意义(P>0.05);Ⅲ组中三类螺死亡率两两比较亦无差异(P>0.05)。
2.4 感染率 Ⅰ组三类螺均无感染;Ⅱ组中,黑色福寿螺、黄色福寿螺、灰色福寿螺的感染率分别为48.15%(13/27)、25%(7/28)、33.33%(8/24),两两比较无统计学意义(P>0.05);Ⅲ组中,黑色福寿螺、黄色福寿螺、灰色福寿螺感染率分别为61.54%(16/26)、40.74%(11/27)、76.19%(16/21),灰色福寿螺感染率高于黄色福寿螺(P<0.05),其余两两比较无差别(P>0.05)。同类螺Ⅱ组与Ⅲ组比较,除灰色福寿螺Ⅲ组感染率明显高于Ⅱ组(P<0.01)外,其余两类螺高剂量感染组与低剂量感染组感染率无差别(P>0.05)。
2.5 感染度及虫密度 Ⅱ组和Ⅲ组3类福寿螺之间感染度及虫密度均无明显差别(P>0.05);同类螺不同组间进行比较,黑色福寿螺和灰色福寿螺Ⅱ组和Ⅲ组间比较感染度和虫密度均无明显差别(P>0.05),黄色福寿螺Ⅱ组和Ⅲ组感染度有差别(P<0.05),Ⅲ组高于Ⅱ组,虫密度无差别(P>0.05)。福寿螺最低感染度出现在Ⅲ组灰色福寿螺,最高感染度出现在Ⅲ组黑色福寿螺(见表2)。将福寿螺感染广州管圆线虫数量分为三个等级(见图3):小于100条、100-500条和大于500条。Ⅱ组无感染大于500条螺存在,小于100条的福寿螺占多数;Ⅲ组中开始出现感染大于500条L3的螺,甚至超过 1 000条。
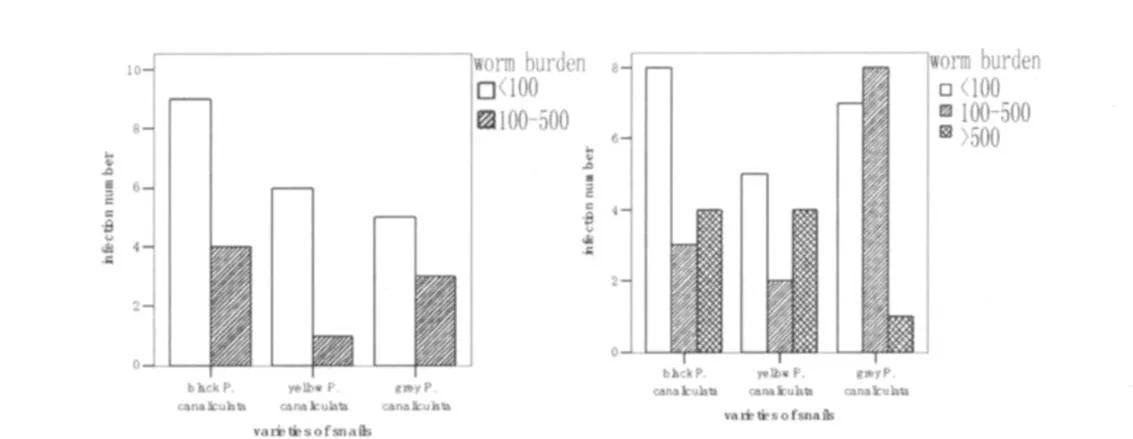
图3a Ⅱ组3类福寿螺的感染情况图3b Ⅲ组3类福寿螺的感染情况Fig.3a Infection status of P.canaliculata in GroupⅡ.Fig.3b Infection status of P.canaliculata in GroupⅢ.
2.6 螺重与易感性关系 对Ⅱ、Ⅲ组三类福寿螺感染广州管圆线虫螺和无感染螺的重量进行比较,发现无差别(P>0.05),即福寿螺是否感染在成年螺中与其大小无关。而在感染L3的螺中,螺的重量与感染度亦无相关性。
3 讨 论
在福寿螺的原发地——南美洲报道福寿螺有多种兼性和专性共生体[13-14],而无福寿螺感染广州管圆线虫报道,也并未对人类健康造成威胁[15]。而在福寿螺入侵地,主要在亚洲,感染广州管圆线虫的福寿螺致人患病已有多次暴发流行,给人的健康造成了严重威胁[5]。我们根据当地福寿螺外部形态将福寿螺分为3类,探讨其对广州管圆线虫易感性,揭示其此生物学特点的不同,并为广州管圆线虫病的防治提供参考。
本实验于春季进行,平均气温超过20℃,且实验过程中供给充足食物,无虫感染时,福寿螺的生长应该是连续的[16],且较小的螺生长更快[17]。然而在本实验中,实验组福寿螺均无生长,说明广州管圆线虫感染可能使螺的生长出现了停滞。实验组螺在感染后活动率下降也说明广州管圆线虫的感染对福寿螺产生了负面影响。黑色福寿螺和灰色福寿螺实验组与阴性对照组比较死亡率无差别,说明螺的死亡并非是由于感染引起,其死亡可能与气温水质、寿命等因素有关[16]。黄色福寿螺阴性对照组死亡大于实验组,由于黄色福寿螺的感染率及感染度均较低,可能小剂量的广州管圆线虫的感染增强了其对外界环境的抵抗力,相关原因还需进一步的研究。
Ⅱ组三类福寿螺感染率及感染度无差别,而在Ⅲ组中,黄色福寿螺感染率低于灰色福寿螺,三类螺感染度无差别。由于灰色福寿螺重量较黑色福寿螺和黄色福寿螺小,我们还对三类螺的虫密度进行了分析,发现同组三类福寿螺亦无差别。本实验中,各感染组福寿螺感染率均较低,感染度亦不高,这与Smarn等[18]人的实验结果相似,在他的研究中,随着感染剂量的增加,虫的回复率无升高甚至出现下降。可见,福寿螺感染广州管圆线虫的个体差异较大,这种差异掩盖了感染剂量不同带来的影响。然而,在本实验中,我们发现实验组即使在未发现L3的福寿螺体内亦有虫鞘膜存在(Ⅱ组中数量较多,Ⅲ组中数量较少),这说明福寿螺被感染过,只是虫由于某些未知因素的作用已死亡并被吸收,这种作用可能与福寿螺自身免疫以及虫之间的相互作用有关[19]。另外,通过对实验组螺重量的分析发现,福寿螺是否感染及感染虫量的多少与螺的重量大小无关。
总之,福寿螺属外来入侵物种,不同的生存环境下会产生不同的变异,包括外部形态的变异和某些生物学特征的变异。本文在对不同品系福寿螺对广州管圆线虫易感性的研究中发现,黄色福寿螺对广州管圆线虫的抵抗性较强。这可能与黄色福寿螺在中国多个省份所占比例较黑色福寿螺多的现状有关[20]。当然,有关福寿螺其他的变异还需要更多的研究来证实。
(感谢武鸿美在福寿螺显微结构的描述上对我们的帮助)
[1]蔡汉雄,陈日中.新的有害生物——大瓶螺[J].广东农业科学,1990(5):36-38.
[2]Nishimura K,Mogi M,Okazawa T,et al.Angiostrongy lus cantonensis infection in Ampullarius canaliculatus(Lamarck)in Kyushu,Japan[J].Southeast Asian J T rop Med Public Health,1986,17:595-600.
[3]Nomura S,Lin PH.First case report of human infection with Haemostrongy lus ratti Yokogawa[J].Taiwan No Ikai,1945,3:589-592.
[4]Kliks MM,Palumbo NE.Eosinophilic meningitis beyond the Pacific Basin:the global dispersal of a peridomestic zoonosis caused by Angiostrongylus cantonensis,the nematode lungworm of rats[J].Soc Sci Med,1992,34:199-212.
[5]Chen XG,Li H,Lun ZR,et al.Angiostrongyliasis,mainland China[J].Emerg Infect Dis,2005,11:1645-1647.
[6]Prociv P,Spratt DM,Carlisle MS,et al.Neuro-angiostrongyliasis:unresolved issues[J].Int J Parasitol,2000,30:1295-1303.
[7]Estebenet AL,Mart n PR.Pomacea canaliculata(Gastropoda:Ampullariidae):Life-history Traits and their Plasticity[J].Biocell,2002,26:83-89.
[8]Cazzanige NJ.Old species and new concepts in the tax onomy of Pomacea(Gastropoda:Ampullariidae)[J].Biocell,2002,26:71-81.
[9]叶嵘,温周瑞,叶奕佐.两种大瓶螺饲养效果对比试验初报[J].渔业致富指南,2000(21):42.
[10]孙志坚,曾清香,张波,等.中国不同品系福寿螺主要形态比较[J].中国人兽共患病学报,2011,27(3):184-189.
[11]Barcante JM,Barcante T A,Dias SR,et al.A method to obtain axenic Angiostrongy lus vasorum first-stage larvae from dog feces[J].Parasitol Res,2003,89:89-93.
[12]Hata H,Kojima S.Angiostrongy lus cantonensis:in vitro cultivation from the first-stage to infective third-stage larvae[J].Exp Parasitol,1990,70:476 482.
[13]Damborenea C,Brusa E,Paola A,et al.Variation in wo rm assemblages associated with Pomacea canaliculata(Caenogastropoda,Ampullariidae)in sites near the R o de la Plata estuary,Argentina[J].Biocell,2006,30:457-68.
[14]Gamarra-Lugues CD,Vege IA,Kock E,et al.Intrahost distribution and transmission of a new species of cyclopoid copepod endosymbiotic to a freshwatersnail,Pomacea canaliculata(Caenogastropoda,Ampullariidae),from Argentina[J].Biocell,2004,28:155-64.
[15]Vega IA,Damborenea MC,Gamarra-Luques C,et al.Facultative and obligate symbiotic associations of Pomacea canaliculata(Caenogastropoda,Ampullariidae)[J].Biocell,2006,30:367-375.
[16]Estebenet AL,Cazzaniga NJ.Growth and demog raphy of Pomaceacanaliculata(Gastropoda:Ampullariidae)under laboratory conditions[J].Malacol Rev,1992,25:1-12.
[17]T anaka K,Watanabe T,Higuchi H,et al.Density dependent growth and reproduction of the apple snail,Pomacea canaliculata:a density manipulation experiment in a paddy field[J].Res Popul Ecol,1999,41:253-262.
[18]Smarn T,Tuanchai S,Paiboon S,et al.Angiostrongylus cantonensis:Experimental study on the susceptibility of apple snails,Pomacea canaliculata compared to Pila polita[J].2008,118:531-535.
[19]Johansson A,Sumpter DJ.From local interactions to population dy namics in site-based models of ecology[J].Theor Popul Biol,2003,64:497-517.
[20]徐建荣,沈辉,李旭光,等.不同品系福寿螺壳色性状的遗传规律分析[J].江苏农业科学,2008(4):210-212.
