茄链格孢菌侵染马铃薯叶片过程的细胞学观察
2011-05-22左豫虎韩青梅康振生台莲梅
左豫虎 史 洁 韩青梅 康振生 台莲梅 刘 铜
(1黑龙江八一农垦大学,黑龙江 大庆 163319;2西北农林科技大学植物保护学院,陕西省农业分子生物学重点实验室,陕西 杨凌 712100)
马铃薯早疫病(Potato early blight)是一种由茄链格孢菌〔Alternaria solani(Ell.et Mart.)Jones et Grout〕引起的对马铃薯危害比较严重的病害,可以导致马铃薯产区大面积减产,带来巨大的经济损失(Pelletier & Fry,1990)。该病在我国各大马铃薯产区均有不同程度发生,局部地区其危害程度不亚于晚疫病(杨殿贤和苑风瑞,2007)。利用组织病理学和电镜技术研究茄链格孢菌与马铃薯互作的组织病理学和超微结构特征,可为在细胞水平上阐明茄链格孢菌的致病机制及马铃薯的抗病机制提供理论依据。Dita等(2007)对马铃薯早疫病病菌侵染过程进行了研究,发现茄链格孢菌可从表皮细胞的相邻部位直接侵入,没有观察到从气孔入侵,并且发现侵染与品种抗性无关,但侵染后抗病品种较感病品种表现出明显的过敏性坏死反应。而在我国鲜见茄链格孢菌与马铃薯互作的组织病理学和细胞学研究的相关报道。本试验采用扫描和透射电镜技术系统研究了茄链格孢菌在抗、感早疫病的马铃薯离体叶片上的侵染过程及超微结构特征,旨在为从细胞水平上阐明茄链格孢菌的致病机制及马铃薯的抗病机制提供理论依据。
1 材料与方法
1.1 供试品种
供试马铃薯品种为东农303(感病)和克新1号(抗病),由黑龙江省克山农场马铃薯研发中心提供。品种抗性由黑龙江八一农垦大学植物病理研究室接种鉴定(台莲梅 等,2010)。
1.2 供试菌株
供试茄链格孢菌菌株SH0806为2008年分离自黑龙江省绥化市采集的马铃薯品种富津(辽宁省本溪市马铃薯研究所培育)的早疫病病叶,由本大学植物免疫研究室提供。。
1.3 分生孢子悬浮液的制备
试验于2009年10月~2010年3月在西北农林科技大学陕西省农业分子生物学重点实验室进行。用打孔器打取PDA平板上生长5~7 d、直径为5 mm的茄链格孢菌菌碟,接入水琼脂平板,25 ℃黑暗培养3 d后产生大量分生孢子,加少量无菌水轻轻震动,使分生孢子能够充分的散落于水中。用血球计数板将分生孢子悬浮液的浓度调整到1.0×105个·mL-1备用。
1.4 离体叶片接种方法
选择3~4叶期的马铃薯同一部位叶片,取1.3制备的孢子悬浮液10 μL,滴于叶片背面,用于透射电子显微镜取样;取孢子悬浮液10 μL滴于叶片正面,用于扫描电子显微镜取样,以蒸馏水接种为对照。置于放有1~2张湿润滤纸的培养皿内,25 ℃恒温培养箱中黑暗保湿培养。
1.5 扫描电镜(SEM)样品的制备与观察
分别于接种后2、4、6、8、10 h取样,将接种部位的叶片切成1 cm2的小块,按康振生(1995)的方法制备样品。叶样经抽真空后置于4 %的戊二醛磷酸缓冲液(0.1 mol·L-1、pH 6.8)中4 ℃固定过夜,然后用相同浓度磷酸缓冲液(PBS)冲洗5~6次,每次20 min,用30 %、50 %、70 %、80 %、90 %、100 %乙醇脱水,每次30 min,再用100 %丙酮脱水2次,每次30 min,最后用醋酸异戊脂置换2次,每次30 min,二氧化碳临界点干燥、粘样、喷金后在JEOL-JSM-6360LV型扫描电镜下观察、拍照。
1.6 透射电镜(TEM)样品的制备与观察
分别于接种后12、24、48、72 h取样,将接种部位的叶片切成0.3~0.5 cm2的小块,按康振生(1995)的方法制备样品。叶样经抽真空后置于4 %戊二醛磷酸缓冲液(0.1 mol·L-1、pH 6.8)中4 ℃固定过夜,然后用相同浓度磷酸缓冲液(PBS)冲洗5~6次,每次20 min,于1 %锇酸磷酸缓冲液(0.1 mol·L-1、pH 6.8)中固定2 h,用30 %、50 %、70 %、80 %、90 %、100 %乙醇脱水,每次30 min,再用100 %丙酮脱水2次,每次30 min,Epon812环氧树脂包埋,在30 ℃和60 ℃下分别聚合24 h和48 h,最后用玻璃刀半薄切片定位,定位后用钻石刀进行超薄切片。半薄切片于光学显微镜下观察侵染点菌丝扩展情况并拍照,超薄切片经醋酸双氧铀—柠檬酸铅双重染色,于JEM-1230型透射电镜下观察、拍照。
2 结果与分析
2.1 分生孢子在叶片上的萌发
扫描电镜观察发现,感病品种东农 303叶片表面未萌发的茄链格孢菌分生孢子形态饱满(图版Ⅰ-1);在适宜培养条件下,接种2 h后,分生孢子开始萌发(图版Ⅰ-2),分生孢子萌发时间在不同抗性品种叶片上无明显差异(图版Ⅰ-2、3、4);分生孢子可从孢子的多个位置(头部、侧面、尾部)萌发(图版Ⅰ-2、3、4),萌发后孢子因营养物质的消耗而凹陷变瘪(图版Ⅰ-3、4);接种6 h后,抗病品种克新1号叶片上的菌丝沿叶片表面蔓延生长,有些菌丝产生了明显的分泌物(图版Ⅰ-5),而有些菌丝没有产生分泌物(图版Ⅰ-6)。

图版Ⅰ 茄链格孢菌分生孢子在马铃薯叶片上萌发的扫描电镜图
2.2 分生孢子在叶片上的侵入
扫描电镜观察发现,接种 6 h后,马铃薯叶片表面茄链格孢菌菌丝大量蔓延,少数菌丝沿着寄主表皮细胞相邻的凹陷处生长(图版Ⅱ-1);随着菌丝在寄主表皮细胞表面的扩展蔓延,菌丝顶端形成附着孢,部分菌丝越过气孔蔓延生长(图版Ⅱ-2),没有观察到通过气孔侵入表皮细胞(图版Ⅱ-2);接种8 h后,菌丝从表皮细胞相邻的凹陷处直接侵入感病品种东农303叶片表皮细胞(图版Ⅱ-3);接种10 h后,菌丝从表皮细胞相邻的凹陷处直接侵入抗病品种克新1号的叶片表皮细胞(图版Ⅱ-4)。说明茄链格孢菌侵入感病品种比侵入抗病品种早2 h。
2.3 菌丝在寄主体内的扩展
透射电镜观察发现,接种12 h后,茄链格孢菌菌丝在感病寄主细胞间隙生长(图版Ⅲ-1);接种24 h后,菌丝穿过感病寄主细胞壁,存在于细胞壁和细胞膜之间,细胞膜变得不连续,可能是病原菌释放出某些物质降解了寄主细胞膜(图版Ⅲ-2),菌丝从一个细胞向另一个细胞生长(图版Ⅲ-3);接种48 h后,大量菌丝存在于感病寄主细胞内和胞间(图版Ⅲ-4)。

图版Ⅱ 茄链格孢菌分生孢子在马铃薯叶片上萌发与侵入的扫描电镜图
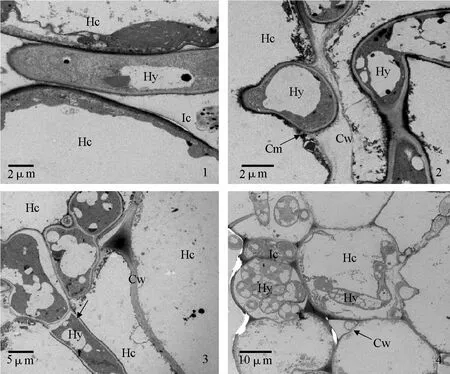
图版Ⅲ 茄链格孢菌菌丝在寄主体内扩展的透射电镜图

图版Ⅳ 寄主细胞病理变化的透射电镜图

图版Ⅴ 寄主细胞病理变化和茄链格孢菌菌丝在不同抗性品种叶片中的侵染情况
2.4 寄主组织的病理变化特征
接种24 h后,抗病品种克新1号寄主细胞壁明显加厚(图版Ⅳ-1),感病品种东农303未观察到细胞壁加厚的现象,细胞内含物已基本全部消解(图版Ⅳ-2);接种48 h后,抗病品种寄主细胞内含物出现不同程度的消解,还可见部分细胞器(图版Ⅳ-3、4)。与感病品种接种24 h后寄主细胞内含物基本全部消解相比,抗病品种细胞消亡速度明显慢于感病品种。
接种 72 h后,抗病品种细胞内含物基本全部消解(图版Ⅴ-1),蒸馏水接种的对照寄主细胞内线粒体、叶绿体等各类细胞器均匀排列,清晰可见(图版Ⅴ-2)。接种 48 h后,光学显微镜观察半薄切片可见感病品种表皮细胞下侵染点存在大量菌丝(图版Ⅴ-3),抗病品种表皮细胞下侵染点的菌丝(图版Ⅴ-4)明显少于感病品种。
3 结论与讨论
利用光学显微镜、扫描和透射电子显微镜观察茄链格孢菌侵染不同抗性马铃薯品种叶片的超微结构特征,发现接种2 h后,分生孢子开始萌发,分生孢子各个部位(头部、侧面、尾部)都能萌发;接种6 h后,菌丝顶端出现附着孢;接种8 h后,菌丝从表皮细胞间隙直接侵入感病品种表皮细胞内,接种10 h后才进入抗病品种表皮细胞内;接种24 h后,感病品种表皮细胞中侵染菌丝向相邻细胞扩展,而此时抗病品种细胞壁出现明显加厚的防卫反应,说明抗病品种对茄链格孢菌的侵染有一定的抗侵入和抗扩展能力。随着茄链格孢菌在寄主细胞内的扩展,叶片发生了一系列的病理变化。接种24 h后,感病品种寄主细胞膜不连续,细胞内含物及各类细胞器已基本全部消解,抗病品种部分消解;接种48 h后,侵入抗、感病品种寄主表皮细胞内的菌丝数量差异明显,感病品种菌丝量明显多于抗病品种,抗病品种细胞内含物此时才基本全部消解。这表明抗病品种对茄链格孢菌的抗性机理存在抗菌丝扩展能力。
本试验中分生孢子萌发、侵入时间明显早于Dita等(2007)报道的接种6 h后分生孢子萌发,接种24 h后菌丝直接侵入感病品种,接种36 h后菌丝直接侵入抗病品种的试验结果,这可能与供试菌株和寄主品种差异或与试验条件有关,其原因有待于进一步研究。本试验中发现抗病品种在接种24 h后出现细胞壁加厚的防卫反应,而感病品种未观察到防卫反应现象。Dita等(2007)报道抗病品种的抗性表现是过敏性坏死反应。这是分别在细胞水平和宏观水平对抗性表现的分析描述,并不矛盾。
病原菌的侵入方式一般分为 3种:伤口、气孔、直接侵入。本试验发现分生孢子萌发与品种抗性无关,部分菌丝在寄主表皮蔓延越过气孔生长,没有观察到菌丝通过气孔侵入寄主的现象,菌丝从表皮细胞间隙直接侵入,这与Dita等(2007)的研究结果一致。本试验方法中,扫描电镜样品在叶片正面接种取样,透射电镜在叶片背面接种取样。原因是:马铃薯属双子叶植物,叶片背面气孔数和绒毛数明显多于正面。正面接种扫描电镜观察,有利于避免绒毛茂密对观察效果的影响;背面接种制备透射样品,若病原菌能够通过气孔侵入,叶片背面气孔数目多则利于茄链格孢菌的侵入。本试验中还发现有些菌丝四周伴随分泌物存在,该类分泌物的存在应该有利于分生孢子在寄主表面的附着与萌发,对其成分和作用有待于进一步研究证实。
康振生.1995.植物病原真菌的超微结构.北京:中国科学技术出版社:5-74.
台莲梅,梁伟伶,左豫虎,金光辉,靳学慧.2010.马铃薯不同品种感染早疫病菌后防御酶活性变化.植物生理学通讯,46(11):1147-1150.
杨殿贤,苑风瑞.2007.25 %嘧菌酯悬浮剂防治马铃薯早疫病田间药效试验.农药科学与管理,28(8):28-29.
Dita M A,Brommonschenkel S H,Matsuoka K,Mizubuti E S G.2007.Histopathological study of the Alternaria solani infection process in potato cultivars with different levels of early blight resistance.Journal of Phytopathology,155(7-8):462-469.
Pelletier J R,Fry W E.1990.Characterization of resistance to early blight in three potato cultivars:receptivity.Phytopathology,80(4):361-366.
