一个支持芥蓝起源于中国的分子证据
2011-05-21王冬梅王庆彪刘玉梅杨丽梅张扬勇方智远
王冬梅 陈 琛,2 王庆彪 庄 木* 刘玉梅 杨丽梅张扬勇 方智远
(1中国农业科学院蔬菜花卉研究所,北京 100081;2青岛农业大学,山东 青岛 266109)
芥蓝(Brassica oleracea var. alboglabra Baily),又名白花芥蓝、芥兰,是十字花科芸薹属一年或两年生草本植物,2n=18,在我国广东、广西、福建和台湾等地广泛种植(Panda et al.,2003;秦耀国 等,2009;张平真,2009;周禹 等,2010)。有关甘蓝类作物起源和亲缘关系的信息多数来自史料记载(Helm,1963;Thompson,1976;张平真,2009):普通结球甘蓝和紫甘蓝大约在13世纪起源于德国;皱叶甘蓝可能原产于意大利;抱子甘蓝形成于比利时的布鲁塞尔;Boswell(1949)认为球茎甘蓝是15世纪在欧洲北部演化而成;花椰菜可能是在1490年由热那亚人从黎凡特或者塞浦路斯引入意大利;青花菜由黎凡特、塞浦路斯或者克里特岛引入意大利;芥蓝的起源有中国南方起源说和地中海沿岸起源说两种,尚缺乏充分的分子生物学证据。
SSR标记由于其多态性好以及共显性的特点(Powell et al.,1996),越来越多地应用于基因标记、图谱构建、遗传多样性分析等方面(陈全求 等,2008)。与基因组SSR相比,EST-SSR作为功能基因的一部分,在功能基因鉴定和种间通用性上更有其优越性(Gupta et al.,2003)。本试验利用甘蓝 EST-SSR标记,对芥蓝、野生甘蓝、观赏羽衣甘蓝、苤蓝、皱叶甘蓝、结球甘蓝、紫甘蓝、抱子甘蓝、青花菜和花椰菜等甘蓝类材料进行遗传多样性分析,比较芥蓝与其他甘蓝类作物之间的亲缘关系,为芥蓝起源提供分子证据。
1 材料与方法
1.1 试验材料
供试材料包括甘蓝类作物自交系、地方品种、一代杂种和野生甘蓝等38份。其中3份野生甘蓝由英国国际园艺研究中心(Warwick HRI,UK)引进,5份芥蓝地方品种由华南农业大学雷建军教授提供,花椰菜由中国农业科学院蔬菜花卉研究所花椰菜课题组提供,其余材料由中国农业科学院蔬菜花卉研究所甘蓝课题组收集、提供(材料编号和名称见表 1)。所有材料于2009、2010年定植于中国农业科学院蔬菜花卉研究所南口实验农场,在芥蓝成熟期调查其植物学性状,调查标准参照《芥蓝种质资源描述规范和数据标准》(李锡香和方智远,2008)。苗期取幼嫩叶片,采用改良CTAB法提取DNA。

表1 供试38份甘蓝类材料名称、类型及来源
1.2 EST-SSR分析
试验所用EST-SSR引物参见陈琛等(2010)的文献,利用8份不同甘蓝变种的材料进行引物初筛,筛选出61对EST-SSR引物,并对所有38份材料进行扩增分析。PCR反应体系为10μL,其中包括:200μmol·L-1dNTPs,上链和下链引物各2.5 pmol,0.5 U Taq酶,1×Buffer(内含1.5mmol·L-1MgCl2),25 ng DNA模板,所用试剂均购自北京博迈德科技发展有限公司。PCR扩增反应条件:94 ℃预变性4min;94 ℃变性30 s,合适温度退火30 s,72 ℃延伸1min,35个循环;72 ℃延伸7min。各引物的合适退火温度根据引物合成报告单而定。8 %聚丙烯酰胺凝胶电泳分离PCR产物,160 V恒压。快速银染法染色。
1.3 数据分析
在相同迁移率位置上,有带记为1,无带记为0,缺失记为999。为保证数据准确、可靠,每块胶板均由两人独立记录,然后比对,有争议的数据作缺失处理。应用NTSYSpc2.11软件采用非加权类平均法(UPGMA)进行聚类分析,得到遗传相似系数,根据遗传相似系数建立聚类图。
2 结果与分析
2.1 EST-SSR扩增分析
本试验利用8个具有代表性的材料对EST-SSR引物多态性进行筛选,共得到61对扩增稳定、条带清晰、品种间带型差异较明显、易于识别的引物。用这61对EST-SSR引物对38份甘蓝类材料扩增,共扩增出174条多态性条带,每对引物扩增的等位位点数不等,变幅为1~5,平均每对引物扩增2.86条带,引物多态性总体较好。其中,引物BoE332在38份材料中的扩增结果见图1。

图1 引物BoE332在部分甘蓝类作物中的SSR扩增结果
2.2 芥蓝与其他甘蓝类作物EST-SSR标记的聚类分析
根据扩增结果建立数据矩阵后用NTSYSpc2.11软件计算不同甘蓝变种间的遗传相似系数,UPGMA法进行聚类分析得到38份甘蓝类材料的聚类分析树状图(图2)。
由图2可知,在相似系数大约为0.62处,所选甘蓝材料按地域分为三大类群:Ⅰ野生甘蓝、观赏羽衣甘蓝、结球甘蓝、紫甘蓝、皱叶甘蓝、苤蓝和抱子甘蓝。这几类除皱叶甘蓝外,其他的在史料记载中均起源于欧洲西北部沿海地区。Ⅱ青花菜和花椰菜。起源于地中海沿岸。Ⅲ芥蓝。首先作为一个单独的类群聚出,表明其与前两大类群的甘蓝类植物在进化途径上有所不同,推测其起源于中国南方。
在相似系数约为0.77处,13份芥蓝材料可分为三类(图2)。东升芥蓝(38)是一高代自交系,聚为第一类。桃山中花(31)可叶、薹兼用,本地铁种(32)为叶用芥蓝,均表现为叶形椭圆、叶面平滑,聚为第二类。其余为第三类,这类材料除长叶中熟(35)叶形为椭圆外,其他材料特点是叶形宽卵圆,叶面皱缩;老种香菇(34)、长叶中熟为薹用类型,其中长叶中熟的主薹为紫红色,侧薹商品性好;日本绿(36)来自于日本,与老种香菇、长叶中熟的亲缘关系较近;香港迟花(37)来自香港,与矮脚筷(33)的亲缘关系较近。芥蓝的植物学性状见表2。
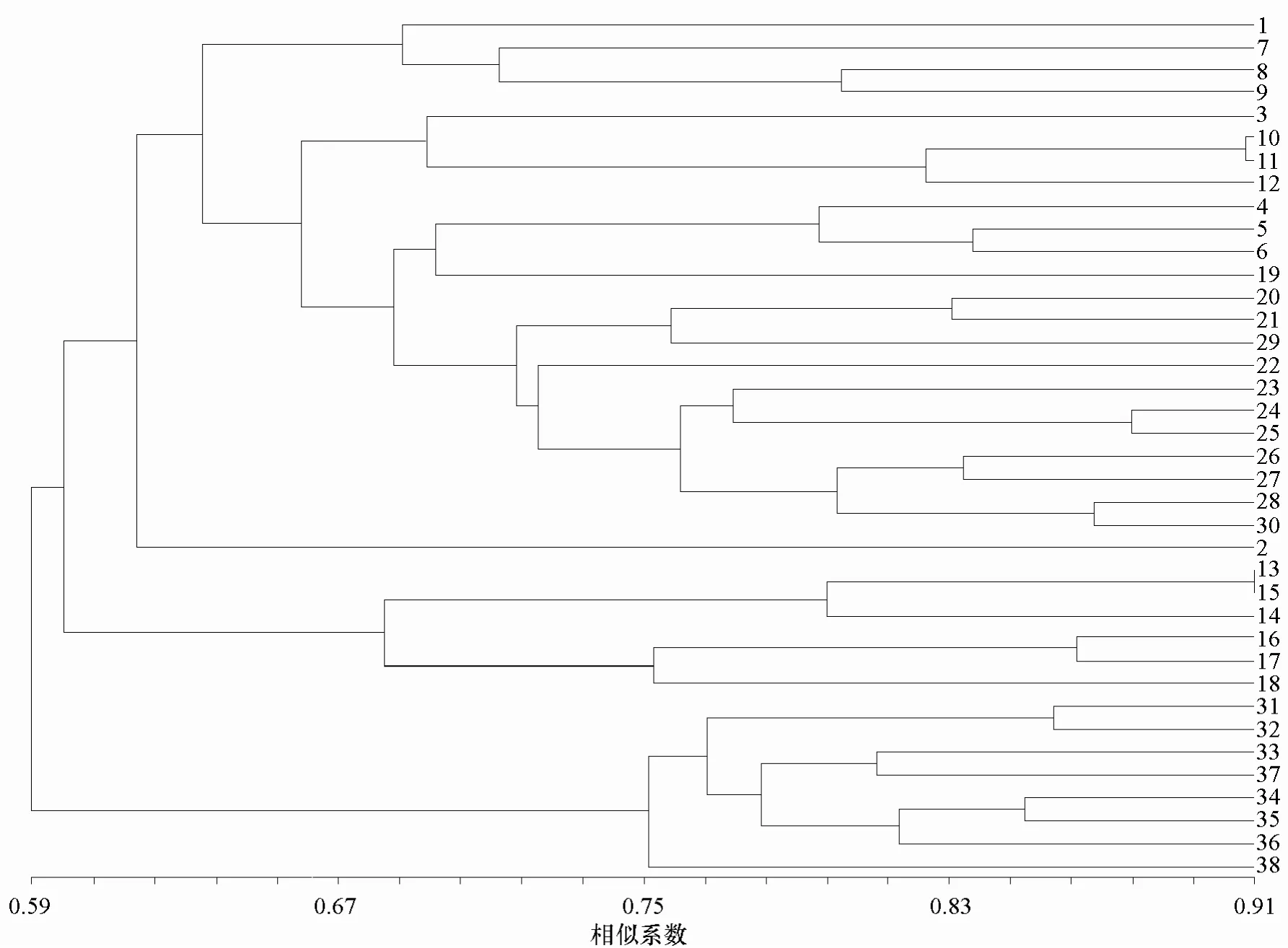
图2 基于EST-SSR分析的38份甘蓝类材料聚类树状图
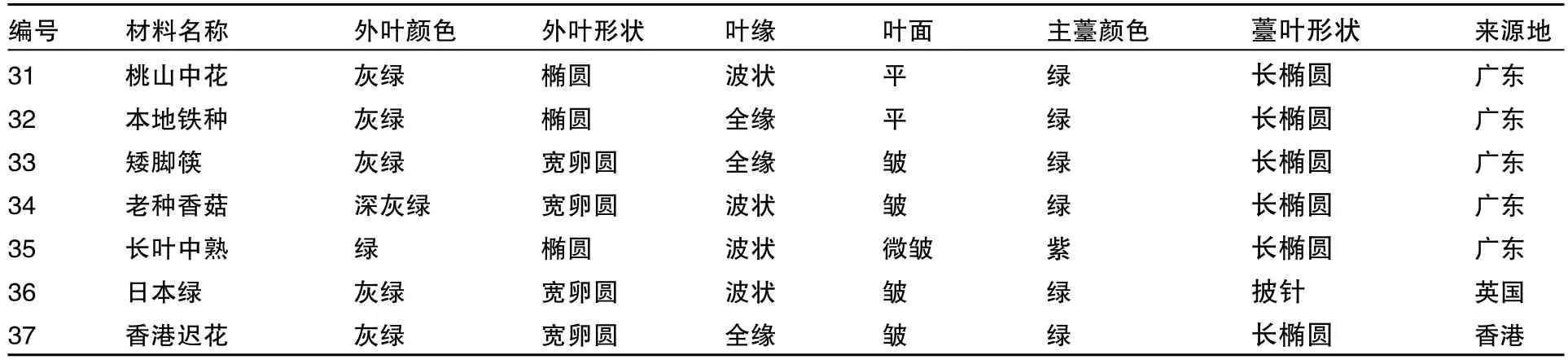
表2 芥蓝植物学性状调查结果
3 讨论
本试验将芥蓝分为三类,除高代自交系东升芥蓝独为一类外,其他两类与Okuda和Fujime(1996)根据植物学性状的分类结果及Matsui等(2002)的RAPD分析结果基本一致,主要是根据叶形和叶面进行分类。
芥蓝的起源现存有两种观点,一是中国南方起源说,在中国目前能追溯到的最早有关芥蓝的记载是宋朝苏东坡(1037~1101)的诗句“芥兰如菌蕈,脆美牙颊响”,距今已有 900多年的历史;而《中国蔬菜栽培学》(中国农业科学院蔬菜花卉研究所,1987)记载最早引进中国的结球甘蓝和花椰菜,也只有300余年的历史,说明芥蓝可能原产于中国;张平真(2009)综合了各种史学资料认为芥蓝为甘蓝的一个变种,并支持芥蓝起源于中国南方的说法。二是地中海沿岸地区起源说,支持这一学说代表著作有《植物学》(中山大学和南京大学,1984)和《中国的蔬菜》(章厚朴,1988)。本试验中芥蓝首先作为一个单独的类群与其他甘蓝类作物分开,表明芥蓝与其他甘蓝类作物可能是由不同路线进化而来。野生甘蓝、观赏羽衣甘蓝、皱叶甘蓝、结球甘蓝、紫甘蓝、苤蓝、抱子甘蓝起源于欧洲西北部沿海地区;青花菜和花椰菜起源于地中海沿岸;芥蓝独立于欧洲西北部类群和地中海沿岸类群,推测其可能起源于中国。有记载早在公元前5、6世纪,一种被称为“西土蓝”的甘蓝类作物传入中国(张平真,2006),在我国副热带气候条件下,逐渐选育形成芥蓝;Song等(1988)的 RFLP研究中芥蓝为单独一类,与千头羽衣甘蓝亲缘关系较近,被认为是一种较为原始的类型,可能起源于一种由西欧传到东亚的原始羽衣甘蓝;本试验中芥蓝作为一个单独的类群首先聚出,与其他甘蓝材料的亲缘关系较远,可能是芥蓝进化形成时间较早,且进化形成的地域与其他甘蓝类作物隔离,支持芥蓝起源于中国的说法。本试验中选用的野生甘蓝都属于Brassica oleracea,全部来自英国,均聚于欧洲西北部沿海类群,表明羽衣甘蓝、皱叶甘蓝、结球甘蓝、紫甘蓝、苤蓝、抱子甘蓝可能都起源于野生甘蓝(Brassica oleracea);而至于芥蓝的祖先,还需进一步收集范围更广的野生甘蓝和近缘野生二倍体进行分析。
陈琛,庄木,李康宁,刘玉梅,杨丽梅,张扬勇,程斐,孙培田,方智远.2010.甘蓝EST-SSR标记的开发与应用.园艺学报,37(2):211-228.
陈全求,詹先进,蓝家样,黄云.2008.EST分子标记开发研究进展.农业生物技术科学,24(9):72-77.
李锡香,方智远.2008.芥蓝种质资源描述规范和数据标准.北京:中国农业出版社.
秦耀国,杨翠芹,曹必好,陈国菊,雷建军.2009.芥蓝遗传育种与生物技术研究进展.中国农学通报,25(18):296-299.
章厚朴.1988.中国的蔬菜.北京:人民出版社.
张平真.2006.中国蔬菜名称考释:甘蓝类蔬菜.北京:北京燕山出版社.
张平真.2009.关于芥蓝起源的研究.中国蔬菜,(14):62-65.
中国农业科学院蔬菜花卉研究所.1987.中国蔬菜栽培学.北京:农业出版社.
中山大学,南京大学.1984.植物学.北京:人民教育出版社.
周禹,李燕,孙勃,石瑜,汪俏梅,汪炳良.2010.芥蓝与甘蓝其他变种分类关系的研究.园艺学报,37(7):1161-1168.
Boswell V R.1949.Our vegetable travelers.Natl Geogr Mag,96:145-217.
Gupta P K,Rustgi S,Sharma S,Singh R,Kumar N,Balyan H S.2003.Transferable EST-SSR marker for the study of polymorphism and genetic diversity in bread wheat.Mor Gen Genomics,270:315-323.
Helm J.1963.Morphologisch-taxonomische Gliederung der kultursippen von Brassica oleracea.Kulturplfanzen,11:92-210.
Matsui T,Okuda N,Kosugi Y.2002.Classification of Chinese kale cultivars by RAPD analysis.Journal of the Japanese Society for Horticultural Science,71(4):499-503.
Okuda N,Fujime Y.1996.Plant growth characters of Chinese kale (Brassica oleracea L. var. alboglabra).Acta Horticulturae,407:55-59.
Panda S,Martin J P,Aguinagalde I.2003.Chloroplast and nuclear DNA studies in a few members of the Brassica oleracea L. group using PCR-RFLP and ISSR-PCR markers:a population genetic analysis.Theor Appl Genet,106(6):1122-1128.
Powell W,Machray G C,Provan J.1996.Polymorphism revealed by simple sequence repeats.Trends Plant Sci,7(2):215-222.
Song K M,Osborn T C,Williams pH.1988.Brassica taxonomy based on nuclear restriction fragment length polymorphisms(RFLPs).1.Genome evolution of diploid and amphidiploid species.Theor Appl Genet,75:784-794.
Thompson K E.1976.Cabbage,kale,etc//Simmonds N W.Evolution of crop plants.New York:Longman:49-52.
