加拿大一枝黄花幼苗生长对土壤水分、光强的响应
2011-05-09徐燕云吴晓梅麻汉林郭水良
徐燕云,吴晓梅,麻汉林,郭水良
(1. 丽水学院生态学院,浙江 丽水 323000;2. 上海师范大学生命与环境科学学院,上海 200234)
加拿大一枝黄花幼苗生长对土壤水分、光强的响应
徐燕云1,吴晓梅1,麻汉林1,郭水良2*
(1. 丽水学院生态学院,浙江 丽水 323000;2. 上海师范大学生命与环境科学学院,上海 200234)
设定4个不同水分和光照梯度,测定加拿大一枝黄花的比叶质量、根生物量比、茎生物量比、叶生物量比、叶质量分数、根冠比来评估其对土壤水分、光强的响应。结果显示,土壤相对含水量60% ~ 90%,中等光照强度(45%)最适合加拿大一枝黄花的生长。
加拿大一枝黄花;水分;光照强度
自然界中植物的生境条件在时间和空间上都存在着很大差异[1],水分和光照是植物赖以生存也是影响其生长的重要生态因子[2~3]。形态特征和生长特性的改变是植物适应不同环境和资源水平的重要策略[4],表型可塑性不仅使物种具有更宽的生态幅和更好的耐受性,占据更加广阔的地理范围和多样化的生境,即成为生态位理论中的广幅种,还可以缓冲甚至在一定程度上屏蔽新生境造成的选择压力,使得入侵种群不至于因大量个体被淘汰而导致种群数量急剧下降。由于表型可塑性和适应性联系密切,人们很早就提出表型可塑性可能是某些入侵物种的重要特征[5]。
王俊峰[4]研究过紫茎泽兰(Eupatorium adenophorum)、兰花菊三七(Gynurasp.)形态特征对不同光照强度的响应,得出两种入侵植物都能很好的适应变幅较大(相对光照强度12.5% ~ 100%)的光环境,许凯扬等[6~7]研究了入侵植物喜旱莲子草(Alternanthera philoxeroides)对不同光照强度、土壤水分的形态可塑性,表明光照强度和土壤水分是影响其入侵的重要环境因子。研究入侵植物加拿大一枝黄花在不同水分及光照条件下的形态适应性,可以为防治加拿大一枝黄花和对不同生境的可入侵性评估作一定的理论依据。
1 材料与方法
1.1 试验设置
本试验在浙江师范大学生物园内进行,加拿大一枝黄花幼苗为采自植物园内的克隆苗,高4 ~ 5 cm,叶片数5 ~ 6片,移栽于高20 cm、内径30 cm的塑料盆中,每盆中装土15 kg,为了防止盆内外水分交流,在盆外垫一层塑料薄膜。金华地区气候温和,四季分明,属亚热带季风气候,年平均温度17.3℃,极端低温-9.8℃,极端高温41.3℃,年降水量为1 500 ~ 1 800 mm。
1.2 水分处理
利用塑料薄膜遮挡自然降水,薄膜透光率为80%。土壤水分处理设置4个梯度:W1为盆内水位高于土壤表面2 cm;W2为土壤水分保持在田间最大持水量的90%以上,每天早晚浇透水;W3为土壤水分保持在田间持水量的60%左右;W4为土壤水分保持在田间持水量的30%左右。每个处理3盆,随机区组排列。2007年3月15日将幼苗移栽于盆内,幼苗成活后备用,于4月1日开始水分处理。
1.3 光照处理
利用黑色塑料遮阳网遮阴,建立相对光照强度分别为100%(不遮阴)、45%、15%和5%的荫棚4个。2007年3月20日将幼苗移栽于盆内,置100%光照下,成活后备用,于4月1日将盆移至各荫棚内。
1.4 测量指标及方法
试验处理后每隔一周用精度为1 mm的直尺测定植株高度,并记录叶片数(为完全展开的叶片)。5月18日和6月6日分别将水分处理、光照处理整株植物小心从盆内挖出,在自来水下冲净泥土带回实验室后分离根、茎、叶,随机挑选10片叶片(成熟叶片),测量其长和宽,叶面积采用“长×宽×0.65”计算,并称质量。将根、茎、叶放80℃烘箱中烘12 h至恒质量,用电子天平(精度为0.001 g)称质量。计算如下参数:比叶质量(Leaf mass per area,叶质量/叶面积)、根生物量比(Root mass ratio,根质量/植株总质量)、茎生物量比(Stem mass ratio,茎质量/植株总质量)、叶生物量比(Leaf mass ratio,叶质量/植株总质量)、叶质量分数(Leaf mass,叶质量/植株地上部分质量)、根冠比(Root /Crown ratio,根生物量/地上部分生物量)。
2 结果与分析
2.1 不同土壤含水量和光照强度对加拿大一枝黄花植株株高、叶片数的影响
不同土壤水分和光照强度对加拿大一枝黄花幼苗株高和叶片数的影响见图1、图2,最终株高和叶片数的方差分析见表1。加拿大一枝黄花植株株高和叶片数均随着处理时间的延长而增加,变化趋势也比较一致。加拿大一枝黄花植株株高在不同土壤水分处理下有极显著的差异,其中W3条件下株高最高,平均高度达43.6 cm, W4条件下最低,只为W3的60%;叶片数在不同处理下有显著性差异,W2条件下达到最高为46.4片,W4最低为36.8片,为W2处理的79%。

图1 不同土壤含水量和光照强度对加拿大一枝黄花株高的影响Figure 1 Effects of different soil water content and light intensity on height of S. canadensis

图2 不同土壤含水量和光照强度对加拿大一枝黄花叶片数的影响Figure 2 Effects of different soil water content and light intensity on the leaf number of S. canadensis
2.2 不同土壤含水量和光照强度对加拿大一枝黄花植株叶面积、比叶质量的影响

表1 株高和叶片数方差分析Table 1 ANOVA of height and leaves
从图3中可以看出,随着土壤含水量的减少,加拿大一枝黄花的平均叶面积呈增加趋势,在W3条件下达到最大值16.9 cm2,而后开始下降。不同处理之间有极显著差异,其中W1条件下平均叶面积只有8 cm2,仅为W3条件的47.3%。在光照强度处理中,不同处理之间有极显著差异,当光照强度在5% ~ 45%时,加拿大一枝黄花平均叶面积随着光照强度的增加而增加,在45%光照下达到最大值15 cm2,随着光照强度的进一步增加,叶面积开始下降,在5%光照强度下叶面积最小,只有45%光照下的一半,而100%光照下亦只达最大值的74%。
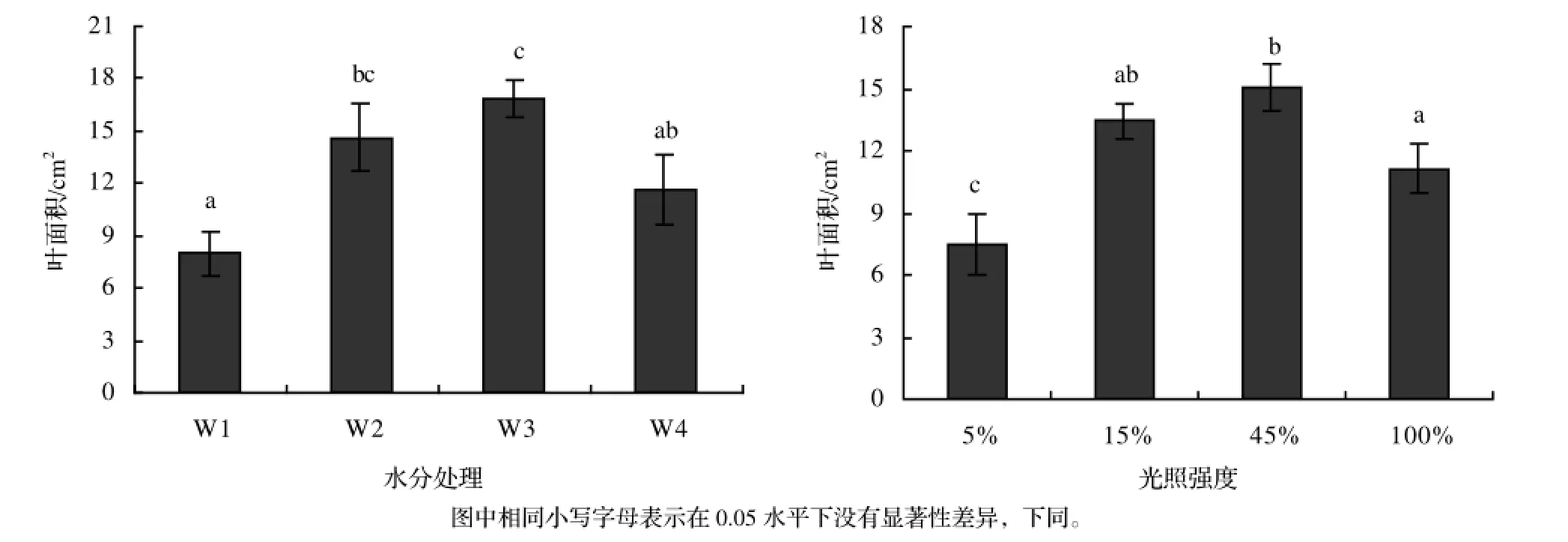
图3 不同土壤含水量和光照强度对加拿大一枝黄花叶面积的影响Figure 3 Effects of different soil water content and light intensity on leaf area of S. canadensis
土壤水分含量对加拿大一枝黄花叶片的比叶质量无明显影响,各处理之间无显著性差异(图4),最低值(W4)和最大值(W3)之间只相差10%。而在光照强度处理中,加拿大一枝黄花叶片比叶质量随光照强度的增加而增加,不同处理之间有显著性差异,5%光照强度处理下比叶质量最低,只有最高(100%光照强度)的61%,说明光照强度对加拿大一枝黄花的比叶质量影响较大。
2.3 不同土壤含水量和光照强度对加拿大一枝黄花植株生物量及生物量分配的影响
2.3.1 土壤含水量变化对加拿大一枝黄花植株生物量及生物量分配的影响 从W1(水生)到土壤相对含水量为60%(W3)时,加拿大一枝黄花的生物量呈增加趋势,而后随着土壤含水量的降低,加拿大一枝黄花的生物量降低(表2),不同处理之间达到极显著的差异。
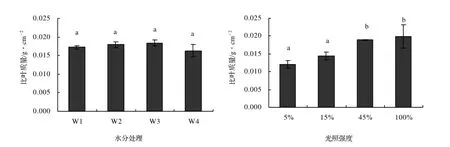
图4 不同土壤含水量和光照强度对加拿大一枝黄花比叶质量的影响Figure 4 Effects of different soil water content and light intensity on leaf mass per area of S. canadensis
根生物量比,根冠比随着土壤水分的减少而逐渐降低,但是在W4条件下又有所增加,各处理间无显著性差异。茎生物量比呈倒“S”型变化趋势,在W1条件下达到最高,各处理间有显著性差异。叶生物量比、叶质量分数在W2条件达到最高,随着土壤相对含水量的减少,其变化趋势呈开口向下的抛物线,各处理间分别达极显著和显著性差异。
2.3.2 光照强度对加拿大一枝黄花植株生物量及生物量分配的影响在相对光照强度5%~45%的范围内,加拿大一枝黄花植株生物量随着光照强度的增加而增加,光照进一步增强时,生物量反而下降,45%光照强度下生物量达到最高,各处理间有极显著差异。
根生物量比、根冠比在各处理间表现出极显著差异,变化趋势呈开口向上的抛物线,极低(5%)和极高(100%)的光照强度均增加了根生物量。中等光照强度(15%,45%)下,加拿大一枝黄花的茎生物量比极显著的高于极端光照强度(5%,100%)下的茎生物量比。叶生物量比、叶质量分数均在5%光照强度下达到最大值,其他几个光照强度下差别较小,处理间有极显著差异。

表2 不同土壤含水量处理下加拿大一枝黄花植株的生物量分配Table 2 Biomass distribution of S. canadensis under different soil water content

表3 不同光照强度处理下加拿大一枝黄花植株的生物量分配Table 3 Biomass distribution of S.canadensis under different light intensity
3 结论与讨论
3.1 水分对加拿大一枝黄花的影响
近年来,对植物水分关系的探讨已成为植物生理生态学研究领域的重要课题之一[8]。水分胁迫是制约植物生长发育的主要逆境因子[9],分析物种在不同水分条件下的形态、结构和生殖分配等方面的可塑性差异,可以从中了解该物种对不同水分梯度环境的适应机制[10],加拿大一枝黄花原产北美,主要分布于湿生生境,而加拿大一枝黄花在我国也主要分布于东部沿海地区[11],这些地区雨量充沛,适合加拿大一枝黄花对生境的要求。
生长行为是植物受多种环境因子综合作用的最终表现[12],许多研究表明,植物在不同环境条件下的资源分配格局反映了植物发育对环境的响应规律和资源分配对策[13~14]。加拿大一枝黄花株高、叶片数及生物量受到土壤水分变化的显著影响。在旱生条件下的株高及叶片数均为四个水分处理中的最低值,说明干旱对加拿大一枝黄花的生长极为不利,但是,旱生条件下植株的根生物量比及根冠比却高于中生和湿生条件下植株,地下生物量的增加有利于植株在更大范围内吸收水分,从而缓解干旱胁迫,使植株的生物量得到增加,这可从旱生条件生物量大于水生条件得到证明。这些和何维明[15],关保华[16],许凯扬[6]等人的研究结果相一致。
水生条件是加拿大一枝黄花生长的特例(目前在自然条件下还没有发现水生种群),但是在整个实验期间,加拿大一枝黄花仍能很好的生长。根生物量比,茎(支持结构)生物量比和根冠比与其他处理相比都有较大的增加,叶面积和生物量比有较大的下降,这可能是加拿大一枝黄花对极端水分状况下作出的一种反应,作为一种陆生植物,加拿大一枝黄花没有水生植物那么发达的通气组织,一旦到了水生生境,为了维持植株的生命延续,植株通过增加根系生物量来弥补通气组织欠缺。同时,根系的增加也有利于植株的固着,而茎生物量的增加也提高了植株的抗倒伏能力,水生条件下植株的叶面积最小,更减少了植株对风的阻力,这些都有利于植株更好的生存。
实验结果还显示,虽然在极端水分条件下(如W1,W4),加拿大一枝黄花的生长受到抑制,但是只要能维持其生命,一旦抑制条件解除,加拿大一枝黄花就能快速的生长,显示其顽强的生命力。
3.2 光照对加拿大一枝黄花的影响
生物生境的基本属性之一是异质性。光是最重要的生态因子,作为植物生存、生长和繁殖所必需的资源之一,其在水平空间内的分布是斑块性的[1],植物通过调整其形态及生物量的分配来适应不同的光照环境,对于物种的生存具有重要的意义。王俊峰[4],许凯扬[6],罗学刚[17]分别研究过紫茎泽兰,喜旱莲子草,蛇莓在不同光照环境下植株的生物量分配,生长特征,得出光照极大的影响植株的形态及其生物量分配。
本次实验中,加拿大一枝黄花的株高受到光照的显著影响,在100%光照强度下植株最低,只有最高(15%光照)情况下的53%,但是植株的叶片数在不同处理之间无明显差别,这是植株对不同光照强度的一种生存对策,在高光照条件下,植株光合作用所需光能相对过剩,较矮的株型及较密的叶片,可形成相互遮挡,避免了叶片灼伤。而在低光照条件下,植株可以通过增加株高来获得更多空间的光照。植株的叶面积在光照强度(5% ~ 45%)范围逐渐增加,随着光强的进一步增加叶面积却有所下降,但是,比叶质量却随着光强的增加而增加,研究表明,叶片大而薄,比叶质量小更有利于植物在弱光环境中生存[18~19],而叶面积小、比叶质量大是植物对强光生境的一种适应[20~21]。在100%的光照强度下,加拿大一枝黄花的根生物量和根冠比均比其他光照强度下的大,在高光照条件下,叶片的气孔数相对较多[22],蒸腾作用较大,植株必须通过扩大根系来吸收更多的水分以维持体内水分平衡。
加拿大一枝黄花生物量在不同光照强度下有极显著的差异,在45%光照下达到最大,显然高光照也影响了植株生物量的积累,但是极低光照(5%)更不利于植株的生存,5%光照下的生物量不到最高值的8%,而且植株柔弱,茎干中空,根据实验后期的继续观察,5%光照下的加拿大一枝黄花逐渐死亡。
3.3 建议
根据本次研究,土壤相对含水量60% ~ 90%,中等光照强度(45%)最适合加拿大一枝黄花的生长,但是,在其他土壤水分条件及光照条件下(除了5%),加拿大一枝黄花也能存活,即使是5%光照条件下,加拿大一枝黄花也能存活两个月之久,说明加拿大一枝黄花对极端生境具有较强的耐受性,滩涂、湿地、林下等各种生境都有可能成为加拿大一枝黄花的入侵对象,因此也要做好这些生境的监测工作。
[1] 董鸣. 异质性生境中的植物克隆生长:风险分摊[J]. 植物生态学报,1996,20(6):543-548.
[2] 何炎红,郭连生,田有亮. 白刺叶不同水分状况下光合速率及其叶绿素荧光特性的研究[J]. 西北植物学报,2005,25(11):2 226-2 233.
[3] 郑丽,冯玉龙. 紫茎泽兰叶片化感作用对10种草本植物种子萌发和幼苗生长的影响[J]. 生态学报,2005,25(10):2 782-2 787.
[4] 王俊峰,冯玉龙. 光强对两种入侵植物生物量分配, 叶片形态和相对生长速率的影响[J]. 植物生态学报,2004,28(6):781-786.
[5] 史刚荣,马成仓. 外来植物成功入侵的生物学特征[J]. 应用生态学报,2006,17(4):727-732.
[6] 许凯扬,叶万辉,李静. 入侵种喜莲子草对土壤水分的表型可塑性反应[J]. 华中师范大学学报(自然科学版),2005,39(1):100-103.
[7] 许凯扬,叶万辉,李国民,等. 入侵种喜旱莲子草对光照强度的表型可塑性反应[J]. 武汉植物学研究,2005,23(6):560-563.
[8] 李洪建,柴宝峰,王孟本. 北京杨水分生理生态特性研究[J]. 生态学报,2000,20(3):417-422.
[9] 郭卫华,李波,黄永梅,等. 不同程度的水分胁迫对中间锦鸡儿幼苗气体交换特征的影响[J]. 生态学报,2004,24(12):2 716-2 722.
[10] Stevens K J, Peterson R L, Stephenson G R. Morphological and anatomical responses of Lythrum salicaria L.(purple loosestrife) to an imposed water gradient [J]. Int J Plant Sci,1997(158):172-183.
[11]董梅,陆建忠,张文驹,等. 加拿大一枝黄花——一种正在迅速扩张的外来入侵植物[J]. 植物分类学报,2006,44(1):72-85.
[12] Beadle C L. Growth analysis[A]. Hall E O, J M O Scurlock, H R Bolhar-Norderkampfeds. Photosynthesis and production in a changing environment[M]. A field and laboratory manual, London: Chapman & Hall.1993.
[13]Geber M A,W atson M A,de Kroon H. Morphological plasticity in clonal plants:the foraging concept reconsidered[J]. J Ecol,1997(83):143-152.
[14]Smith N S. Variation in responses to defoliation between populations of Boteloua curtipendula var. eaespitosa with different livestock grazing histories[J]. Am J Bot,1998(85):1 266-1 272.
[15]何维明. 水分因素对沙地柏实生苗水分和生长特征的影响[J]. 植物生态学报,2001,25(1):11-16
[16] 关保华,葛滢,樊梅英,等. 华荠宁响应不同土壤水分的表型可塑性[J]. 生态学报,2003,23(2):259-263.
[17] 罗学刚,董鸣. 蛇莓克隆构型对光照强度的可塑性反应[J]. 植物生态学报,2001,25(4):494-497.
[18] Urbas P. Adaptive and inevitable morphological of three herbaceous species in a multi-species community: field experiment with manipulated nutrients and light[J]. Acta Oecologic,2000(21):139-147.
[19]Lee D W,R A Bone,S Tersis,et al. Correlates of leaf optical properties in tropical forest sun and extreme-shade plants[J]. Am J Bot,1990(77):370-380.
[20]蔡永立,宋永昌. 浙江天童常绿阔叶林藤本植物的适应生态学I. 叶片解剖特征的比较[J]. 植物生态学报,2001,25(1):90-98.
[21] Mendes M M, Gazarini L C, Rodrigues M L. Acclimation of Myrtus communis to contrasting Mediterranean light environments effects on structure and chemical composition of foliage and plant water relations[J]. Environ Exp Bot,2001(4):165-178.
[22]蔡志全,齐欣,曹坤芳. 七种热带雨林树苗叶片气孔特征及其可塑性对不同光照强度的响应[J]. 应用生态学报,2004,15(2):20l-204.
Response of Seedlings Growth of Solidago canadensis to Water and Light Intensity
XU Yan-yun1,WU Xiao-mei1,MA Han-lin1,GUO Shui-liang2*
(1. Ecological College of Lishui University, Lishui 323000, China; 2. College of Life and Environment Science, Shanghai Normal University, Shanghai 200234, China)
The index of leaf mass per area, root mass ratio, stem mass ratio, leaf mass ratio, leaf mass, root /crown ratio of Solidago canadensis seedlings were determined in the different conditions of water and light intensity to evaluate its response to water and light intensity. The result showed that the growth of S. canadensis was the best when the relative soil water content was 60%-90% and the light intensity was 45% .
Solidago canadensis; water content; light intensity
S718.51
A
1001-3776(2011)02-0013-06
2010-11-18;
2011-02-07
浙江省教育厅项目“加拿大一枝黄花入侵生物学及其控制”(200601564)
徐燕云(1971-),女,浙江丽水人,副教授,硕士,从事植物多样性及植物生理生态学研究;*通讯作者。
