退化羊草群落恢复演替系列种群构件年龄结构特征
2011-04-25孟令宇李海燕杨允菲
孟令宇,李海燕,杨允菲
(东北师范大学草地科学研究所 吉林省生态恢复与生态系统管理重点实验室植被生态科学教育部重点实验室,吉林 长春 130024)
羊草(Leymuschinensis)是典型的无性系根茎禾草,营养繁殖力强,在我国内蒙古东部和东北西部以及黄土高原地区的天然草地有广泛分布。羊草具有很高的营养价值,是我国优良的牧草之一。羊草草地是优良的放牧地和割草地,多年来一直是草地学和生态学工作者研究的热点,并已开展到生物学的各个层次和水平[1-5]。以往对羊草的研究已涉及到羊草个体生物生态学特性、种群生物生态学特性和群落生态特性等方面[6]。但对不同演替系列羊草种群构件年龄结构特征的研究尚未见报道。
种群年龄结构是种群内不同年龄个体数量的分布情况,种群年龄结构不仅反映种群的不同年龄个体的组配情况,也反映种群与环境间的相互关系以及种群的发展潜势,对反映种群动态和种群特征都有重要的意义[7]。在天然植被中,许多无性系植物具有旺盛的营养繁殖习性,这种习性往往使得一个无性系种群在局部地区繁荣,并且在有些情况下可保持和繁衍几个世纪[8-13],甚至更长的时间[14]。事实上,许多无性系植物的生理寿命是没有极限的,它们已经找到了永生的秘诀[15]。开展无性系种群年龄结构的研究,不仅可以了解其种群的现在甚至可以预测其未来,也可以从无性系小株出生、死亡的周转中揭示无性系植物寿命的长短以及长寿的秘密。
植物群落的演替是指在植物群落发展变化的过程中,由低级到高级、由简单到复杂、一个阶段接着一个阶段、一个群落代替一个群落的自然演变现象[16]。演替过程中的植被变化可进一步划分为演替阶段,因此可以根据演替阶段来判断植物群落退化或者恢复的情况[17-21]。在草原恢复演替的过程中群落及其种群的变化机理将为退化生态系统的恢复工作奠定理论基础。本研究通过对退化羊草草甸恢复期不同演替系列羊草种群构件的年龄结构特征进行分析,以揭示空间演替过程中羊草种群的发展变化趋势,为松嫩平原退化草原的恢复管理提供科学依据。
1 材料与方法
1.1样地概况 本研究在松嫩平原南部的吉林省长岭种马场、东北师范大学草地生态研究站(44°45′ N, 123°45′ E)[22-23]进行。该地区冬寒少雪,春旱风大,夏暖多雨,属于温带大陆性半湿润季风气候。年平均降水量为435 mm,主要集中在6-8月,约占全年降水量的60%;年蒸发量1 368 mm,约为降水量的3倍;大于10℃的年积温在2 579~3 144℃·d 之间;绝对最高气温可达37.8℃ ,绝对最低气温可达-33.9℃ ,土层结冻深度可达1 m 以上;无霜期136~163 d[22,24]。
研究样地选择因重度放牧已退化的羊草草甸。自然景观为碱斑连片,羊草仅少量分布。该研究样地于1999年开始围栏禁牧封育,在10余年的自然封育中,形成了羊草与主要伴生种比例不同的恢复演替系列。其中,羊草和全叶马兰(Kalimerisintegrifolia)是典型的恢复演替系列之一。群落常见的其他的伴生种有蔓委陵菜(Potentillaflagellaris)、寸草苔(Carexduriuscula)、箭头唐松草(Thalictrumsimplex)、细叶苦荬菜(Ixeridiumgracile)、拂子茅 (Calamagrostisepigeios) 、芦苇(Phragmitesaustralis)、问荆(Equisetumarvense)等。
1.2研究方法 本研究依据羊草密度从低到高划分3个演替系列: 1)全叶马兰+羊草群落,以全叶马兰为优势种,羊草为亚优势种的混生群落(以下简称系列1); 2)羊草+全叶马兰群落,以羊草为优势种,全叶马兰为亚优势种的混生群落(以下简称系列2);3)羊草群落,以羊草单优势群落(以下简称系列3)。2009年10月初,在割草场和围栏封育2个不同处理的样地,以羊草比例从低到高的每个演替系列中取样。取样面积为25 cm×25 cm。每个演替系列5次重复。将每个样方的羊草分株连同地下根茎一并挖出,按样方编号、分别装袋,带回室内逐样方逐个分株划分年龄级别。把分蘖节繁殖再生的世代数看作分蘖植株的年龄,并按形态特征进行鉴别。具体划分及识别方法已做报道[25]。
1.3数据统计与分析 研究结果中,各参数均换算成1 m×1 m的常规单位面积。统计分析中,用平均数(M)代表样本各性状整体水平特征,用标准差(SD)反映样本的绝对变异度。由各个龄级分蘖植株或芽的数量与全体龄级分蘖植株或芽的数量百分比,构成分蘖植株或芽的年龄谱。用Excel和SPSS统计分析软件对数据进行分析。不同演替系列均数采用方差分析(One-way ANOVA) 进行比较,方差齐者两两比较采用Duncan检验,方差不齐者用Tamhane’s T2 检验,显著水平为0.05。
2 结果与分析
2.1分蘖株的年龄结构 研究结果显示,不同演替系列的羊草种群分蘖株存在一定差异(表1)。围栏样地中,系列1由1龄和2龄两个龄级组成,随着演替系列的增加,系列2和3出现了3龄级,在不同演替系列中只有系列2中1龄分蘖株的数量明显高于其他两个系列,不同龄级分蘖株3个演替系列并没有共同的变化规律。围栏样地中不同演替系列的分蘖株的年龄结构特征均是1龄>2龄>3龄,为增长型的年龄结构。割草样地中,系列1和系列3均由2个龄级组成,系列2为3个龄级,不同演替系列的分蘖株的年龄结构特征均是1龄>2龄>3龄,为增长型的年龄结构。割草样地中1龄级分蘖株随着演替系列的增加明显地增加,而2龄、3龄不同演替系列之间差异不显著。在围栏和割草样地的3个演替系列中,平均都以1龄比例占绝对的优势达81.0%和69.1%,2龄明显减少为16.9%和29.7%,3龄仅占极小的比例为2.1%和1.3%。以上结果表明,不同演替系列羊草种群分蘖株均属于增长型的年龄结构。
2.2根茎长度和年龄结构 根茎不仅是重要的繁殖器官,也是营养物质的重要存储器官。羊草根茎累积长度的数量结构既反映羊草的无性繁殖能力又反映其空间扩展能力。不同演替系列的羊草种群根茎长度和年龄结构之间存在显著差异(表2)。围栏样地中不同演替系列根茎总的累积长度没有明显差异,而在割草样地中,系列1明显低于系列2和系列3。围栏样地中根茎长度均以2龄根茎占绝对优势,1龄次之,3、4龄依次呈不同幅度的减少。割草样地中仍然以2龄占绝对的优势,3龄和1龄次之,4龄只占极小的比例。在不同的演替系列中,4龄级的老龄根茎所占比例极小,说明老龄根茎的扩展能力已经极低。在围栏样地中,根茎长度属于增长型的年龄结构,而在割草样地中属于稳定型的年龄结构。1龄的根茎为当年形成的新根茎,围栏样地中新根茎形成的长度要略大于割草样地。围栏与割草样地不同演替系列根茎长度的差异和共同规律还需进一步研究。
2.3芽库存量和年龄结构 生长季末期10月是营养繁殖芽生产的旺盛时期,羊草种群将通过营养繁殖芽进行种群的补充和更新。各龄分蘖节横向生长的根茎顶端芽和根茎节间芽为0龄芽,由1龄分蘖节产生的向上生长的芽为1龄芽,由2龄分蘖节产生的向上生长的芽为2龄芽,以此类推[25]。
围栏和割草样地不同演替系列芽库存量存在一定差异(表3),围栏中系列1与系列2和3存在明显的差异,系列2与系列3无差异。
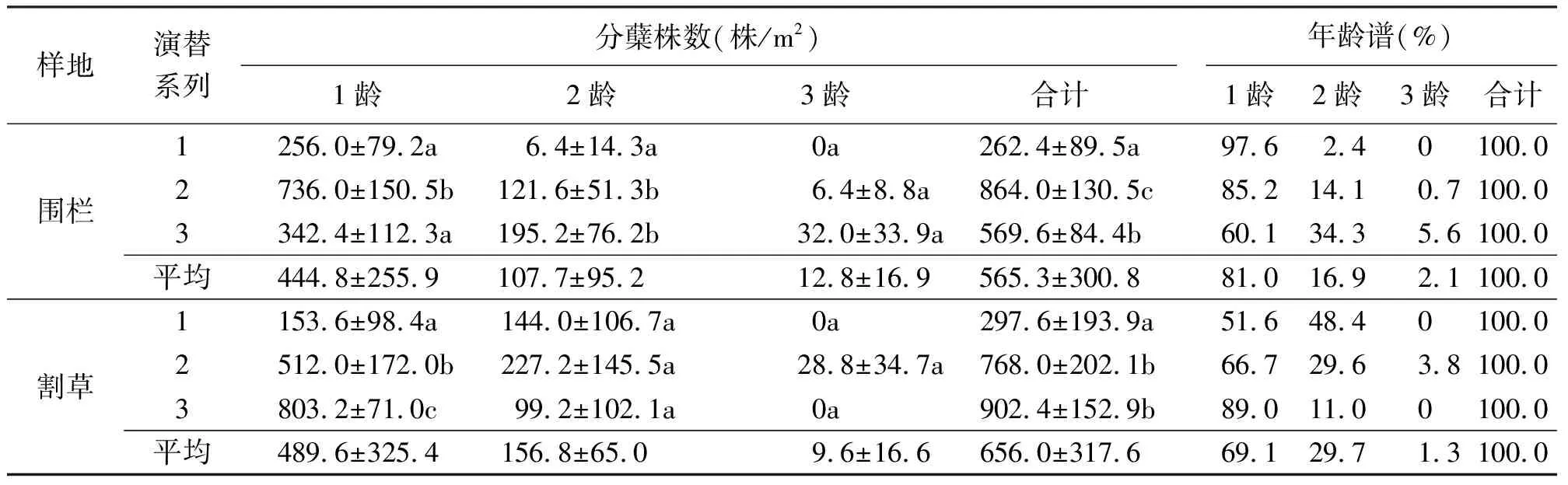
表1 不同演替系列羊草种群分蘖株的年龄结构及年龄谱
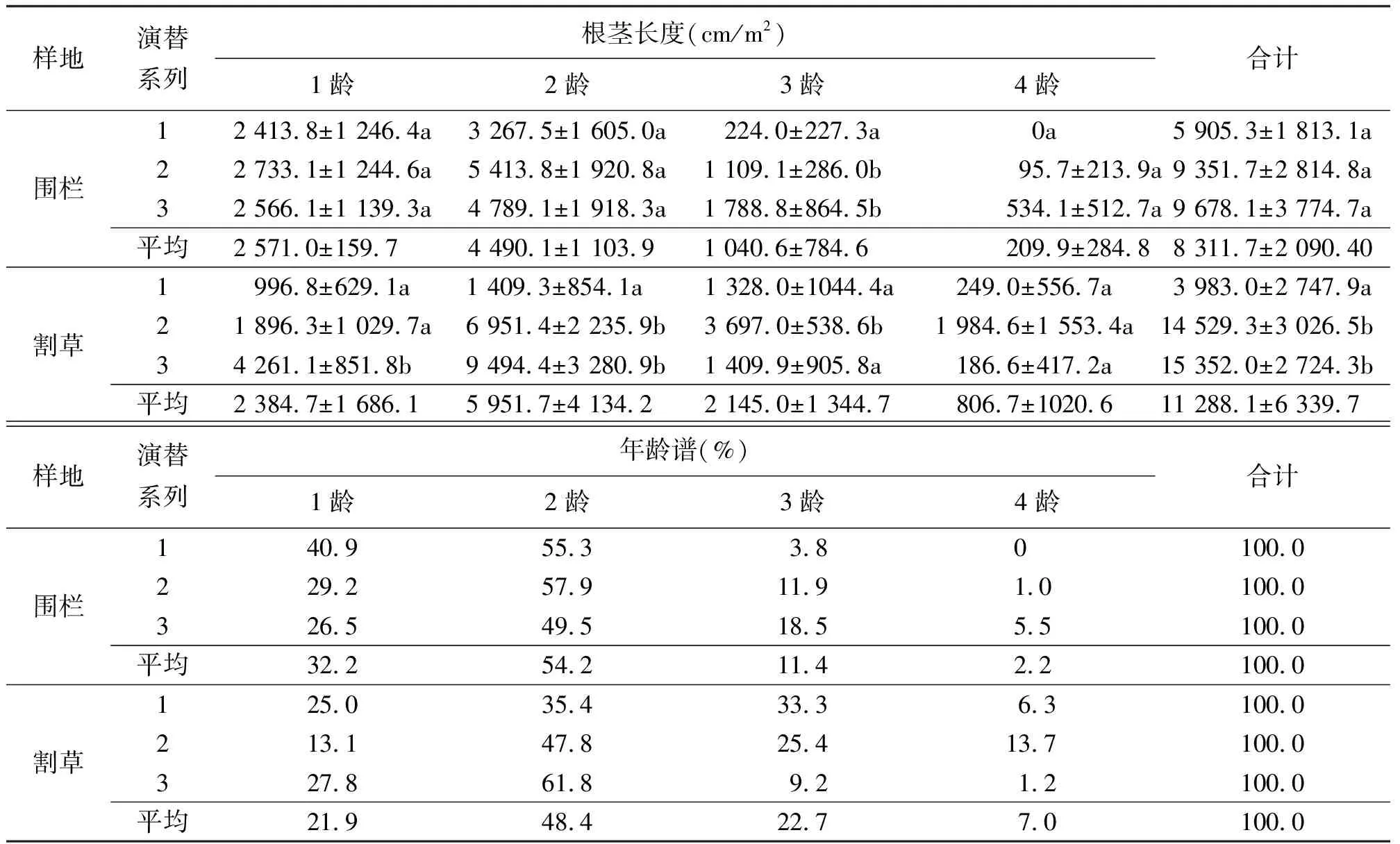
表2 不同演替系列羊草种群根茎长度的年龄结构和年龄谱
而割草样地中,系列1与系列2、系列3存在显著的差异。各样地不同演替系列由4个龄级组成,均以0龄芽数量占优势,随着龄级增加,芽数量呈不同程度减少。由于0龄芽将形成翌年的1龄分蘖株,1龄芽形成2龄分蘖株,依次类推[26],各样地芽库的年龄谱均为0龄>1龄>2龄>3龄为增长型年龄结构。预测翌年的现实种群的结构也为增长型年龄结构。割草场0龄芽的平均值为(1 046.4±773.3)个/m2,而围栏样地中0龄芽的数量是(888.5±219.9)个/m2,表明割草可以促进0龄级芽即根茎芽的形成。两样地间存在的规律需进一步研究。
2.4羊草种群冬眠构件的组成 多年生的草本植物均依靠冬眠构件度过寒冷的冬季,羊草的冬眠构件有两种,一种是以芽的形式冬眠,一种是以莲座丛状分蘖苗的形式冬眠。冬性苗和芽的存量可视为翌年的潜在种群。在生长季的末期,羊草种群已经出现冬性苗和芽两种形式的冬眠构件。
围栏和割草样地中不同演替系列均以根茎芽所占的比例最多(表4),平均达到61.8%和46.6%,其次是分蘖节芽为31.0%和23.2%,冬性苗在围栏中仅占到7.2%,而割草样地中冬性苗平均所占比例达30.2%,反映出割草能极大地促进芽输出形成冬性苗,同时分蘖节芽的数量围栏>割草,根茎芽的数量割草>围栏,表明割草可以促进根茎芽的形成。已有研究表明[26],冬性苗的越冬性能大于芽的性能,在翌年春天冬性苗将形成冬性植株补充种群。由羊草潜在种群的组成可知,羊草根茎的营养繁殖力要大于分蘖节的营养繁殖力,割草可促进冬性苗的形成,在翌年的种群中割草样地羊草种群将出现更多的冬性植株,围栏样地将产生更多的春性植株。围栏中随着演替系列的增加,冬性苗呈明显增加的趋势,分蘖节芽系列1与系列2和系列3之间存在显著性差异,根茎芽系列2与系列1和系列3之间没有显著的差异。
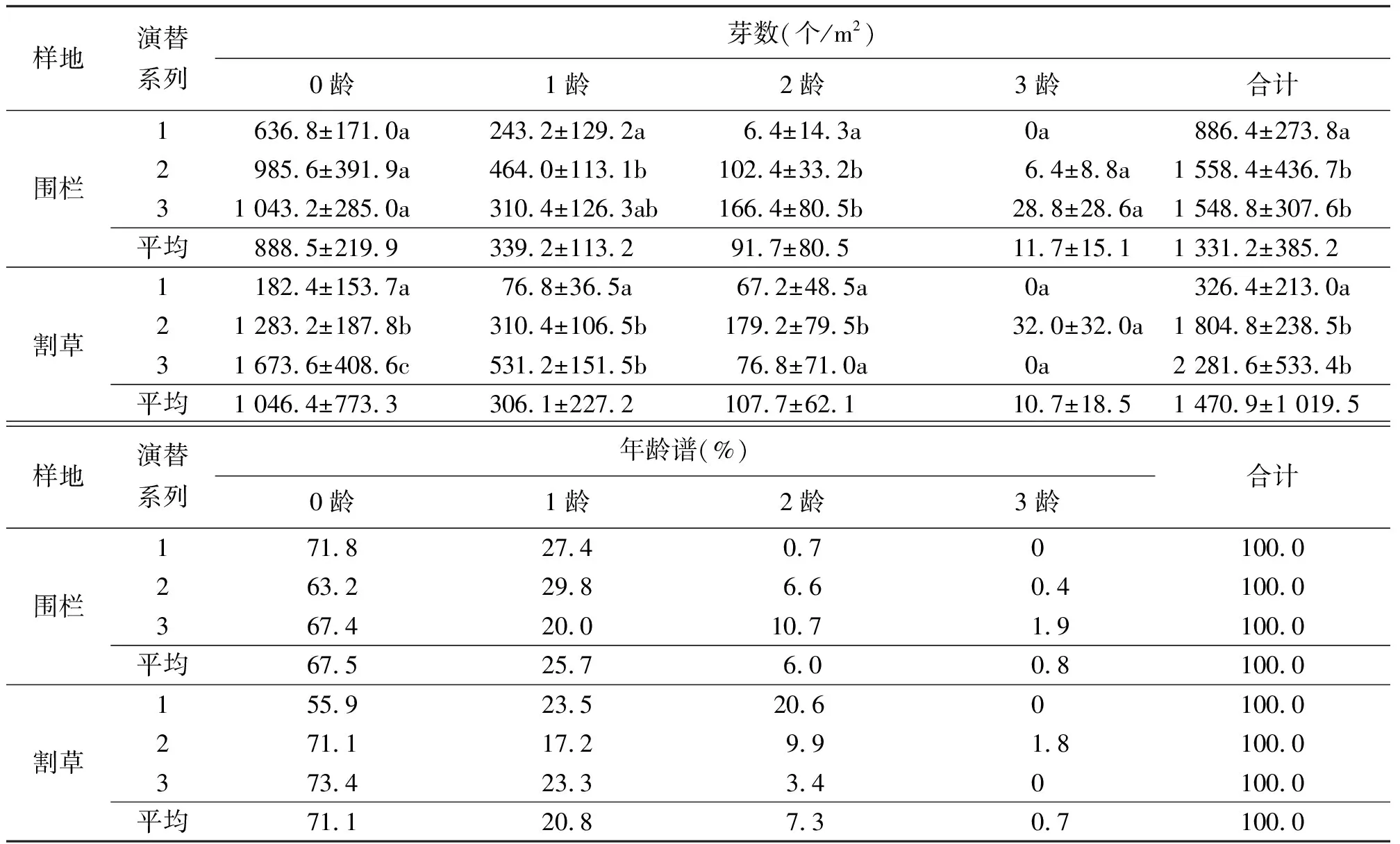
表3 不同演替系列羊草种群芽库存量和年龄谱
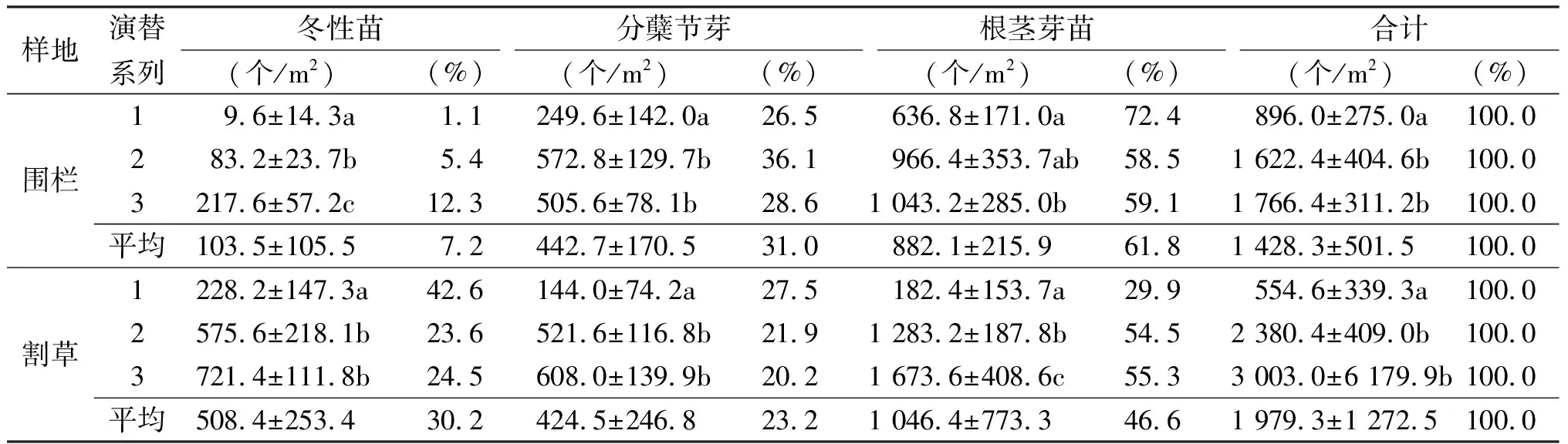
表4 不同演替系列羊草种群冬眠构件存量及比率
3 讨论与结论
羊草是典型的无性系根茎禾草,对羊草分株年龄结构、世代组成、克隆繁殖的研究将揭示种群繁殖的基本规律和基本原理,为人类创造丰产植被与舒适生存空间提供巨大的帮助[27]。目前,人类干扰是草原生态系统退化的原因之一,人类干扰将影响草原植物的多样性及草原植物群落的结构。退化草原中各植物种群对放牧,刈割等人为干扰都有其独特的响应机制,对退化草原恢复演替过程中植物种群的自组织机理的研究是目前生态学的研究热点之一[28-29]。
松嫩平原退化羊草群落围栏与割草草地不同的演替系列中羊草种群分蘖株的年龄结构组成均表现出相同的组成结构,不同演替系列的分蘖株的年龄结构,均是1龄>2龄>3龄。不同演替系列羊草无性系种群分蘖株均属于增长型的年龄结构。不同的演替系列存在一定的差异但规律性不明显,随着群落演替的不断进行,羊草种群空间格局的变化需要进一步的研究。
羊草根茎的累积长度的数量结构既反映羊草的无性繁殖能力又反映其空间扩展能力。不同演替系列羊草根茎均由4个龄级组成,以2龄的根茎所占比例最大,4龄根茎所占的比例最小,反映出2龄根茎对种群的营养繁殖和羊草种群的空间扩展贡献最大,4龄老龄根茎的生命力已经极弱。在围栏样地中,根茎长度属于增长型的年龄结构,而在割草样地中属于稳定型的年龄结构。1龄的根茎为当年形成的新根茎,围栏样地中新根茎形成的长度要略大于割草样地。结合分蘖株的年龄结构分析可以看出羊草地下根茎的数量配置与分蘖株的数量配置有很好的吻合,两者的年龄结构配置具有一致性。
在松嫩平原10月是羊草营养繁殖芽生产的旺盛时期,羊草种群将通过营养繁殖芽进行种群的补充和更新。各样地均由4个龄级组成,均以0龄芽数量占优势,各样地芽库的年龄谱均为0龄>1龄>2龄>3龄呈增长型结构,预测翌年的种群仍为增长型结构。在一定的演替阶段内羊草表现出极强的营养繁殖能力,保持种群呈增长型的年龄结构。禾草在减轻放牧实施围栏后,在自身的强大竞争优势下,将抑制其他植物的生长和扩展,逐渐成为群落的优势植物[30]。因此,随着演替时间的增加羊草将成为松嫩平原的优势种。同时割草可以促进0龄级芽即根茎芽的形成。芽库中各龄级的芽在条件适宜时,最终都可能长出分蘖株。芽的发生及生长状况预示着下一生长季无性系的兴衰,随着演替时间的增加羊草芽库存量的变化规律还需更深一步的研究。
在生长季的末期,羊草种群已经出现两种形式的冬眠构件。羊草可以通过芽和冬性苗两种形式越冬。不同演替系列羊草种群冬眠构件的组成显示出割草干扰可以促进芽的输出形成冬性苗,同时促进根茎芽的形成。由潜在种群预测翌年的羊草现实种群,羊草根茎的营养繁殖力要大于分蘖节的营养繁殖力,割草可促进冬性苗的形成,表明在翌年的种群中割草样地将出现更多的冬性植株,围栏样地中将产生更多的春性植株,两样地种群的植株类型不同。对于潜在无性系构件组分的命运及其在无性系更新与发展中的地位和作用等,有待于进一步研究。
[1] 王昱生.羊草(Leymuschinensis)种群无性系种群动态的初步研究[J].生态学报,1993,13(4):291-299.
[2] 王昱生,李景信.羊草种群无性系生长格局的研究[J].植物生态学与地植物学学报,1992,16(3):234-242.
[3] 王昱生.中国东北羊草草原羊草种群生态的研究Ⅰ.羊草种群数量的初步研究[J].中国草原,1985(3):11-15.
[4] 杨允菲,张宝田.松嫩平原羊草种群营养繁殖的季节动态及其生物量与密度关系的分析[J].植物学报,1992,34(6):443-449.
[5] 包青海,音陶格涛,阎巧玲,等.羊草草原割草处理群落特征比较研究[J].内蒙古大学学报(自然科学版),2003,34(1):74-78.
[6] 祝廷成.羊草生物生态学[M].吉林:吉林科学技术出版社,2004:90-103.
[7] 杨允菲,刘庚长,张宝田.羊草种群年龄结构及无性繁殖对策的分析[J].植物学报,1995,37(2):147-152.
[8] Harberd D J.Observations on population structure and longevity ofFestucarubraL.[J].New Phytologist,1961,60:184-206.
[9] Harberd D J.Some observations on natural clones inFestucaovina[J].New Phytologist,1962,61:85-100.
[10] Harberd D J.Observations on natural clones ofTrifoliumrepensL.[J].New Phytologist,1963,62:198-204.
[11] Cook R E.Clonal plant population[J].American Scientist,1983,71:244-253.
[12] Oinonen E.The Correlation between the size of Finnish brachen [Pteridiumaquilinum(L.)Kuhn)] clones and certain periods of site history[J].Acta Forestalia Fennica,1967,83:1-51.
[13] Oinonen E.Sporal regeneration of ground pine (LycopodiumcomplanatumL.) in southern Finland in the light of the size and age of its clones[J].Acta Forestalia Fennica,1967,83:76-85.
[14] Oinonen E.The time-table of vegetative spreading of the lily-of-the-valley (ConvallariamajalisL.) and the wood small-reed [Calamagrostisepigeios(L.)Roth] in southern Finland[J].Acta Forestalia Fennica,1969,97:1-35.
[15] Silvertown J W.植物种群生态学导论[M].祝宁等,译.哈尔滨:东北林业大学出版社,1987:170-187.
[16] 牛翠娟,娄安如,孙儒泳,等.基础生态学[M].第二版.北京:高等教育出版社,2002:165.
[17] 程积民,邹厚远.黄土高原草地合理利用与草地植被演替过程的试验研究[J].草业学报,1995,4(4):17-22.
[18] 王炜,刘钟龄,郝敦元,等.内蒙古草原恢复演替研究Ⅰ.退化草原的基本特征与恢复动力[J].植物生态学报,1996,20(5):449-459.
[19] 郝郭元,刘钟龄,王炜,等.内蒙古草原退化群落恢复演替的研究——群落演替教学模型[J].植物生态学报,1997,21(6):503-511.
[20] 郑慧莹,李建东.松嫩平原盐生植物与盐碱化草地的恢复[M].北京:科学出版社,1999:84-119.
[21] 杨允菲,郑慧莹.松嫩平原碱斑进展演替实验群落的比较分析[J].植物生态学报,1998,22(3):214-221.
[22] 杨允菲,祝廷成.不同生态条件下羊草种群种子生产的探讨[J].生态学报,1988,8(3):256-262.
[23] 杨允菲,祝廷成.羊草种群种子生产的初步探究[J].植物生态学与地植物学报,1989,13(1):73-78.
[24] 杨允菲,张宝田.松嫩平原羊草种群营养繁殖的季节动态及其生物量与密度关系的分析[J].植物学报, 1992,34(6):266-271.
[25] 杨允菲,郑慧莹,李建东.根茎禾草无性系种群年龄结构的研究方法[J].东北师大学报自然科学版,1998(1):49-53.
[26] 杨允菲,祝玲.松嫩平原十五种多年生禾草种群营养繁殖体冬眠特性的分析[J].草业学报,1994,3(6):26-31.
[27] 洪锐民,王昱生,黄大明.概论植物种群克隆遗传——生态学的任务、性质和产生与发展[J].生态学报, 2004(4):865-867.
[28] 郑伟,朱进忠,潘存德.草地植物多样性对人类干扰的多尺度响应[J].草业科学,2009,26(8):72-80.
[29] 金晓明,韩国栋.放牧对草甸草原植物群落结构及多样性的影响[J].草业科学,2010,27(4):7-10.
[30] 李以康,林丽,张法伟,等.嵩草草甸退化和恢复过程中主要牧草演替和地表特征变化[J].草业学报,2010,19(5):179-185.
