布氏杆菌抗原蛋白pbp39基因重组穿梭质粒的构建与鉴定
2011-04-23杨玉莹长江大学动物科学学院湖北荆州434025
钟 蓓,杨玉莹 (长江大学动物科学学院,湖北荆州434025)
张西臣 (吉林大学畜牧兽医学院,吉林长春130062)
顾玉芳,陈振威,多 婷 (长江大学动物科学学院,湖北荆州434025)
布鲁氏菌病是由布氏杆菌 (Brucella abortus)引起的导致感染动物流产、不孕不育、生殖器官病变的人畜共患传染病,我国大部分地区都有此病的存在及流行。近年来,其疫情形势更为严峻,仅2009年发病率同2008年相比即上升25.37%[1],此病成为人类发病数上升速度最快的传染病之一[2]。布氏杆菌是革兰氏阴性胞内寄生菌,人感染此病后需长期服用抗生素,易产生耐药性及副作用。因此消除布鲁氏菌病成为公共健康计划中最重要的目标之一[3]。目前我国广泛使用的疫苗有牛S19、羊Rev.1和牛RB51,这些疫苗的使用对当地的品种都有较好的效果,但现有的疫苗始终存在不安全、不稳定、且不能与野毒株相区分等缺点。因此,应用分子生物学技术研制新型分子标记苗和基因工程苗成为该病疫苗研究的必然趋势。
布氏杆菌胞质结合蛋白 (Periplasmic binding protein,PBP39)作为良好的T细胞抗原,被认为是DNA疫苗的免疫抗原候选分子。本研究将布氏杆菌抗原基因pbp39和标记基因p276克隆入大肠杆菌-分枝杆菌穿梭表达载体pMV361中,构建重组pMV361-pbp39-p276质粒,为布氏杆菌分枝杆菌疫苗的研制提供了前提条件。
1 材料与方法
1.1 质粒和菌株
穿梭表达质粒pMV361(Stover博士构建)由吉林大学张西臣教授惠赠,E.coli DH5α、布氏杆菌、重组克隆质粒PMD 18-T-p276为长江大学动物科学学院动物病原与分子生物学实验室保存,PMD 18-T simple vector为 Takara公司产品。
1.2 主要试剂
TaqDNA聚合酶为Fermentas公司产品;DNA mareker DL2000、DL250+、T4-DNA ligense、凝胶回收试剂盒为上海捷瑞生物公司产品;质粒少量提取试剂盒为Omega公司产品;限制性内切酶EcoR I,Hpa I,Nhe I为Fermentas公司产品。
1.3 引物的设计与合成
根据GenBank提供的序列,使用Primer 5.0分别设计pbp39和p276基因序列的上游引物和下游引物。在pbp39基因的上、下游引物中分别引入EcoR I、H pa I酶切位点。在p276基因的上、下游引物中分别引入Hpa I、Nhe I酶切位点,并分别在NCBI进行Blaster查看引物的特异性,引物序列为:P1:5'-TAGAATTCATGGGCGCCTGTTGCCAATGC-3';P2:5'-GCGTTAACTATTTTGCGGCTTCAACCGCC-3';G1:5'-TACATGTTAACGATTTAGCCGACCAAACAGCA-3';G2:5'-ATGCTAGCTTAATAGGGCCTTATAAGAAGGTC -3'。引物由上海生工生物技术有限公司合成。
1.4 布氏杆菌基因组DNA的提取及纯化
布氏杆菌 B株接种胰蛋白胨斜面培养基,于 37℃培养 2~3d。消化液 (100mmol/L NaCl,10mmol/L T ris-HCl pH 8.0,25mmol/L EDTA pH 8.0,0.5%SDS)清洗细菌,沸水煮10min裂解细菌,加入蛋白酶K至终浓度为200μ g/mL,55℃温育5~6h,其间需振荡EP管。加入 500μ L苯酚∶氯仿∶异戊醇 (25∶24∶1)抽提2次,再加入500μ L氯仿∶异戊醇 (24∶1)抽提1次。水相用1/8上清体积的5mol/L NaCl和2倍体积预冷的无水乙醇沉淀。70%乙醇洗涤沉淀2次,干燥后溶于灭菌ddH2O中。
1.5 目的基因的扩增
分别以提取的布氏杆菌全基因组DNA和PMD 18-T-p276质粒作为模板,扩增全长为1206bp的pbp39基因和276bp的p276基因。反应条件:94℃预变性5min,94℃55s;65℃1min;72℃1min30s,30个循环;72℃延伸10min。
1.6 含pbp39-p276融合基因的重组穿梭载体的构建
用EcoR I,Hpa I对经PCR扩增获得的pbp39基因片段进行双酶切,克隆入经相同内切酶处理的质粒pMV361中,构建重组质粒pMV361-pbp39,双酶切鉴定后送上海生工公司测序。用 Hpa I,Nhe I对经PCR扩增获得的p276基因片段进行双酶切,克隆入经测序鉴定正确的重组质粒pMV361-pbp39中pbp39片段的下游,构建重组质粒pMV361-pbp39-p276,PCR鉴定后送上海生工公司测序。
2 结果与分析
2.1 布氏杆菌pbp39基因和标记p276基因片段的获得
布氏杆菌pbp39基因PCR扩增产物经1%琼脂糖凝胶电泳后,在1200bp处可见明显的扩增产物条带,未见非特异扩增条带,与理论扩增产物长度吻合。测序结果与GenBank报道的序列进行比对碱基序列同源性为99%(图1)。
p276基因PCR扩增产物经2%琼脂糖凝胶电泳后,在270bp处可见明显的扩增产物条带,未见非特异扩增条带,与理论扩增产物长度吻合 (图2)。

图1 pbp39基因的PCR扩增
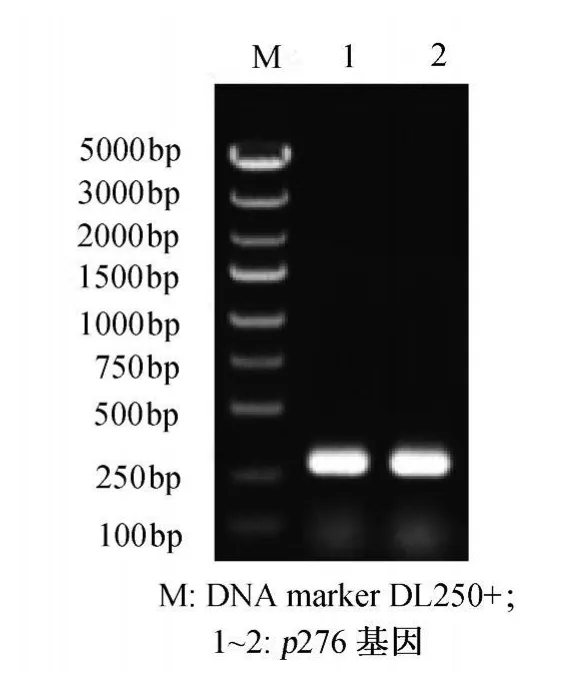
图2 p276基因的PCR扩增
2.2 重组子pMV361-pbp39的鉴定
提取PCR鉴定阳性菌质粒,用EcoR I、Hpa I进行双酶切,得到4415bp的pMV361和1206bp的pbp39 2个片段,基因片段与设计大小相符 (图3)。
2.3 重组质粒pMV361-pbp39-p276的PCR鉴定
提取筛选的阳性菌pMV361-pbp39-p276质粒,分别用 pbp39、p276基因片段的上下游引物,及 pbp39上游引物和p276下游引物进行PCR,扩增得到大小约1200bp、270bp和1500bp的3条带,大小与目的DNA及复合DNA片段长度相符。表明pbp39-p276复合基因已克隆入pMV361载体中 (图 4)。

图3 重组子的酶切鉴定

图4 重组子pMV361-pbp39-p276的PCR扩增鉴定
3 讨论
布氏杆菌为兼性胞内寄生菌,有光滑和粗糙2种表型,主要通过鼻、咽、口腔感染动物。如其他胞内寄生的致病菌,布鲁氏菌不仅能逃避巨噬细胞的杀伤作用,且能在巨噬细胞内复制。宿主只有通过激发抗原递呈细胞分泌IL-12,引起THD细胞分化为THI细胞,THI细胞再分泌IFN-γ来上调巨噬细胞的吞噬作用[4],才能激活巨噬细胞的杀菌功能。同时,CD4+,CD8+T细胞分泌的IL-2,IFN-γ还能直接杀灭被布氏杆菌感染的巨噬细胞,达到保护宿主的目的[5]。因此在抵抗布氏杆菌的免疫反应中,细胞免疫占主导地位。作为保护性抗原分子的PBP39胞质结合蛋白,是一种良好的T细胞抗原,几乎存在于所有的布氏杆菌基因组中,具有较高的同源性,能保证良好的免疫原性,且PBP39在动物体内能诱发强烈的迟发性超敏反应,并产生大量的IFN-γ,因此被公认为DNA疫苗的候选分子。
pMV361大肠杆菌-分枝杆菌穿梭表达载体是在pMV261基础上构建起来的,相对于pMV261,pMV361作为整合型表达载体将其oriM区域替换为attachment site(attP)和integrase(int)gene,使得pMV361能在宿主体内稳定存在[6]。而其表达宿主牛分枝杆菌弱毒苗 (BCG),在表达多种外源基因抗原的同时还是一种免疫佐剂,能非特异性的增加机体的免疫应答,增强免疫效果。且本身作为广泛使用的结核病疫苗,能达到一苗多防的效果,不仅能同时防止布鲁氏菌和结核病的感染,且能区分布鲁氏菌病的疫苗株与野毒株,逐渐达到净化布鲁氏菌的目的。
综上所述,本研究将标记p276基因和抗原pbp39基因联合重组于pMV361大肠杆菌-分枝杆菌穿梭质粒,为下一步构建能分泌性表达pbp39-p276的重组卡介苗基因疫苗,研究其免疫原性和免疫保护性打下了基础。
[1]王大力,李铁锋,王季秋,等.2009年全国布鲁氏菌病监测结果分析 [J].中国地方病防治杂志,2010,25(6):419-421.
[2]满腾飞,王大力,崔步云,等.2009年全国布鲁氏菌病监测数据分析 [J].疾病监测,2010.25(12):944-946.
[3]丁家波,毛开荣,程君生,等.布氏杆菌病疫苗的应用和研究现状[J].微生物学报,2006,46(5):856-859.
[4]Murphy A,Sathiyaseelan J,Parent M A,et al.Interferongamma is crucial for surviving a Brucella abortus infection in both resistant C57BL/6 and susceptible Balb/c mice[J].Immunology,2001,103:511-518.
[5]曾 政,王 英,赵光宇,等.布氏杆菌pCDNA3.1-L7/L12核酸疫苗的构建及免疫学评价[J].免疫学杂志,2004,20(3):208-212.
[6]Stover C K,de la Cruz V F,Fuerst T R,et al.New use of BCG for recombinant vaccines[J].Narure,1991,351:456-460.
