zebularine调控卵巢癌A2780细胞RASSF1A基因再表达的研究
2011-04-19沈文静刘石磊邱广蓉
沈文静,刘石磊,邱广蓉
(1.中国医科大学附属第一医院妇科,辽宁沈阳 110001;2.辽河油田 康复医院,辽宁兴城 125100;3.中国医科大学基础医学院遗传学教研室,辽宁沈阳 110001)
Ras相关区域家族1A基因(Ras associated domain family genes 1A,RASSF1A),是从人3号染色体短臂克隆出来的一种新型肿瘤抑制基因,RASSF1A基因异常甲基化引起RASSF1A表达缺失参与多种人类恶性肿瘤的发生[1]。研究表明,RASSF1A基因异常甲基化导致表达失活与上皮性卵巢癌的发生、发展相关[2]。由于甲基化的过程是可逆的,通过应用去甲基化药物能够诱导由于基因启动子区异常甲基化导致失活的抑癌基因再表达[3],这有可能成为卵巢癌临床治疗的新靶点。zebularine(Zeb)是一种去甲基化药物,具有性质稳定、毒性低的特点[4]。Cheng 等[5]发现,Zeb 具有较强的去甲基化和抗肿瘤增殖作用,提示该药物有可能作为抗肿瘤药物应用于临床。本研究通过观察应用Zeb后人卵巢癌细胞系A2780中抑癌基因RASSF1A甲基化状态及表达水平的改变,探讨Zeb对卵巢癌细胞RASSF1A基因表达的调控作用,初步为临床治疗提供理论依据。
1 材料与方法
1.1 实验材料
人卵巢癌细胞系A2780(天津血液病学研究所),Zeb(Sigma公司),Trizol(Invitrogen公司),RT-PCR 试剂盒及Taq酶(TaKaRa公司),RPMI 1640培养基(Hyclone公司),RASSF1A羊抗人多克隆抗体(Santa Cruz公司),Wizard DNA纯化柱(Promega公司)。
1.2 实验方法
1.2.1 人卵巢癌细胞系A2780的培养条件及实验分组 细胞培养条件:应用含10%胎牛血清的RPMI 1640,培养箱内温度设置 37 ℃,CO2浓度 50 ml·L-1。取对数生长期的A2780细胞,应用含低浓度胎牛血清的培养基饥饿培养24 h,再予实验药物处理,在培养瓶中加入含实验浓度(50、100、200 μmol·L-1)Zeb 的培养液,实验的24、72 h更换同样的加药培养液,于第120小时收集细胞,对照组为未加药物干预的A2780细胞。
1.2.2 RASSF1A基因启动子区甲基化状态的检测应用甲基化特异性 PCR(methylation-specific PCP,MSP)法。实验步骤:(1)离心沉淀收集Zeb作用120 h各实验组A2780细胞,应用酚-氯仿-异戊醇法提取细胞DNA。(2)DNA的修饰及纯化:根据文献已有方法[6]进行。纯化体系 50 μl:样本 DNA 2 μg,3 mol·L-1NaOH 5 μl,充分混匀,加热至 75 ℃,反应 15 min。取 20 mmol·L-1氢醌 12.6 μl(新鲜配置)和 320 μl浓度4.8 mol·L-1的亚硫酸氢钠(NaHSO4),充分混匀,加入矿物油10 μl,55℃水浴过夜。将DNA纯化树脂1 ml加入DNA纯化体系中,混匀充分,再通过纯化柱,应用80%异丙醇2 ml溶解树脂。应用NaOH(3 mol·L-1)脱硫,醋酸钠(5 mol·L-1)及无水冰乙醇沉淀纯化的DNA。应用75%乙醇洗涤后,溶于TE溶液30 μl中备用。(3)MSP反应:设置25 μl反应体系,其中含2 μl样品 DNA,2.5 μl 10 × PCR buffer,0.5 μl引物,2 μl dNTP,0.2 μl Taq 酶,17.3 μl双蒸水。
PCR反应条件为:95℃预变性5 min;94℃ 30 s,58℃ 45 s,72℃ 30 s,35个循环;72℃延伸10 min。退火温度设置:甲基化反应为58℃,非甲基化反应为57℃。阳性对照设置:甲基转移酶SssⅠ处理的正常人外周血细胞DNA;阴性对照设置:未经甲基转移酶SssⅠ处理的正常人外周血细胞DNA;空白对照设置:加入双蒸水。分子质量标记:1 000 bp DL2000。
实验应用的引物序列:甲基化反应,实验目的产物94 bp,上游引物5'-GTGTTAACGCGTTGCGTATC-3',下游引物5'-AACCCCGCGAACTAAAAACGA-3';非甲基化反应,实验目的产物108 bp,上游引物5'-TTTGGT TGGAGTGTGTTAATGTG-3',下游引物5'-CAAACCC CACAAACTAAAAACAA-3'。
PCR产物应用2%琼脂糖凝胶电泳进行分离,电泳条件:电压100 V,电泳时间40 min。电泳结果应用激光密度扫描仪(pharmacia LKB ultroscan)扫描后保存并进行分析。PCR实验重复5次。
1.2.3 RASSF1A 基因 mRNA的检测 应用 RTPCR法。
离心沉淀收集Zeb作用120 h各实验组A2780细胞,细胞总RNA应用Trizol试剂进行提取,定量后稀释为终浓度1 μg·L-1。首先合成 cDNA:应用逆转录酶和Oliga(dT)20作为引物。
RT-PCR反应体系设定为25 μl,反应条件设置为:95℃预变性2 min;94℃ 40 s,67℃ 40 s,72℃60 s,35个循环;72℃延伸6 min。扩增目的产物:RASSF1A mRNA 330 bp;β-actin 498 bp。引物序列设定:RASSF1A mRNA上游引物5'-GGCGTCGTGCGC AAAGGCC-3',下游引物 5'-GGG TGGCTTCTTGCTG GAGGG-3';内参 β-actin上游引物5'-GTGGGGCGC CCCAGGCACCA-3',下游引物 5'-CTCCTTAATGT CACGCACGATTTC-3'。应用2%琼脂糖凝胶电泳对RT-PCR产物进行分离,电泳结果应用自动成像仪(Alpha Image 2000)进行扫描成像并保存,然后采用Fluorchem V 2.0 Stand Alone软件进行分析。RT-PCR实验重复5次。
1.2.4 RASSF1A蛋白表达检测 应用Western blotting法。
离心沉淀收集Zeb作用120 h各实验组A2780细胞。总蛋白提取:应用200 μl蛋白裂解液(细胞 ∶细胞裂解液为1∶5),蛋白定量采用酚试剂法。上样:于总蛋白取样100 μl进行变性反应10 min后,取其中的25 μl加样进行电泳。10%聚丙烯酰胺凝胶电泳:配制10%的SDS聚丙烯酰胺分离胶以及5%浓缩胶,取已变性总蛋白样品20 μl,顺序加入加样孔中,100 V的电压电泳2 h。蛋白质转膜及封闭:将蛋白电泳分离后转移至PVDF膜,应用10%脱脂奶粉进行封闭(室温、1 h)。抗体杂交:然后将PVDF膜放在1∶200一抗稀释液中过夜(4℃),再置于1∶2 000稀释的碱性磷酸酶标记的二抗稀释液中,37℃孵育2 h,DAB显色后观察结果。成像应用ChemiImager 5500 AlPhaInn Ch系统,数据采集应用Fluorchem V 2.0系统。以β-actin蛋白表达为内对照。实验反应重复5次。
2 结 果
2.1 Zeb对A2780细胞中RASSF1A基因甲基化状态的影响
RASSF1A基因在卵巢癌细胞系A2780中呈异常甲基化状态,经过不同浓度 Zeb处理 120 h后,RASSF1A 基因在50 μmol·L-1组甲基化条带减弱,在100、200 μmol·L-1组呈去甲基化状态。见图 1。
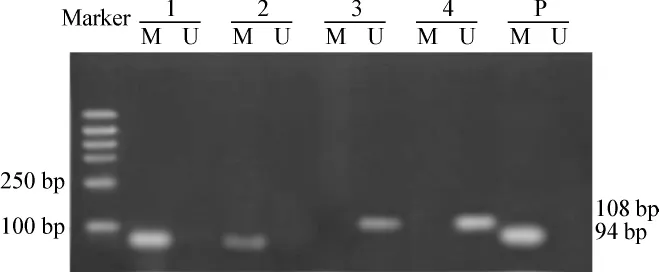
图1 A2780细胞经Zeb作用120 h后RASSF1A基因甲基化状态的改变Fig 1 Change in methylation state of RASSF1A gene in ovarian cancer cell A2780 after treated with Zeb 120 h
2.2 Zeb对A2780细胞中RASSF1A基因mRNA表达的影响
Zeb处理前卵巢癌细胞A2780中未检测到RASSF1A 基因 mRNA 表达,在 100、200 μmol·L-1组Zeb处理120 h后RASSF1A基因在mRNA水平重新表达,见图2。

图2 A2780细胞经Zeb作用120 h后RASSF1A基因mRNA表达的改变Fig 2 mRNA expression of RASSF1A gene in ovarian cancer cell A2780 after treated with Zeb 120 h
2.3 Zeb对A2780细胞中RASSF1A基因蛋白表达的影响
Zeb处理前卵巢癌细胞A2780中未检测到RASSF1A 基因蛋白表达,在 100、200 μmol·L-1组 Zeb处理120 h后RASSF1A基因重新表达,见图3。
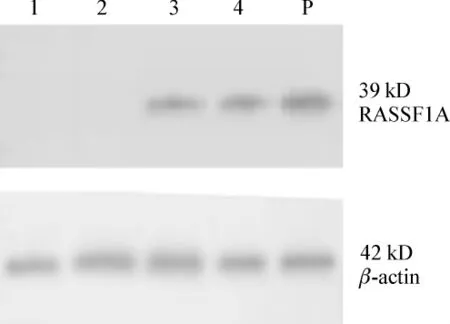
图3 A2780细胞经Zeb作用120 h后RASSF1A基因蛋白表达的改变Fig 3 Protein expression of RASSF1A gene in ovarian cancer cell A2780 after treated with Zeb 120 h
3 讨 论
卵巢恶性肿瘤是女性生殖器三大恶性肿瘤之一,5年生存率25%~30%,其主要原因是缺少有效的早期诊断方法及化疗耐药性。肿瘤抑制基因启动子区异常甲基化导致抑癌基因表达失活参与卵巢癌的发生及发展[2,7]。由于应用去甲基化药物可以使抑癌基因异常甲基化区域恢复到去甲基化状态并恢复功能,目前去甲基化药物的研究正在成为肿瘤治疗领域的热点,可能成为卵巢癌临床治疗的新靶点[4]。相对于目前研究较多的去甲基化药物5-氮-2'-脱氧胞苷(5-aza-2'-deoxycytidine,5-Aza-CdR)存在毒性、不稳定性的缺点[8],并且虽然体外实验对卵巢癌细胞具有生长抑制作用[9],但临床试验对包括卵巢癌在内的实体瘤疗效不理想[10],Zeb具有性质稳定、毒性低的特点,它对肿瘤细胞具有高选择性生长抑制作用,可能用于包括卵巢癌在内的恶性肿瘤的临床治疗[4]。Zeb,[1-(β-D-呋喃核糖苷)-1,2-二氢嘧啶-2-酮],是胞苷类似物,其分子结构同一种已知的去甲基化药物5-氮杂胞苷(5-azacytidine)相似。
RASSF1A基因是一种重要的抑癌基因,RASSF1A蛋白主要以GTP依赖的方式结合Ras传导通路上游的Ras/GTP而被激活,发挥抑制细胞生长、促进细胞凋亡和衰老的功能[11],是Ras传导通路中非常活跃的负向调节蛋白。卵巢癌组织中已证实,RASSF1A基因甲基化与卵巢癌发生有关,异常甲基化是其失活的主要机制[2]。本研究显示,卵巢癌细胞 A2780细胞中RASSF1A基因呈异常甲基化状态,经过 Zeb处理120 h后,RASSF1A 基因在 100 及 200 μmol·L-1组呈现去甲基化,表明Zeb能使RASSF1A基因去甲基化。同时,Zeb处理前A2780细胞中RASSF1A基因在mRNA及蛋白水平均无表达,Zeb处理后,伴随RASSF1A基因去甲基化,其mRNA及蛋白重新表达。根据Zeb处理前后RASSF1A基因甲基化状态及表达的变化,可以看出异常甲基化是卵巢癌细胞RASSF1A基因失活的主要原因,去甲基化药物Zeb通过逆转RASSF1A基因甲基化状态,调控RASSF1A基因重新表达。新近研究[12]表明,结肠癌细胞株HT-29中DLC-1基因表达沉默与启动子区高甲基化状态有关。同时,在急性髓性白血病中RIZ1基因表达下降,其部分机制在于基因启动子区甲基化[13]。这些研究表明,启动子区异常甲基化是抑癌基因失活的主要机制,包括RASSF1A基因。由于RASSF1A能抑制细胞生长、促进细胞凋亡,通过去甲基化诱导RASSF1A抑癌基因重新表达可能是Zeb抑制卵巢癌细胞增殖的原因之一。DNA甲基化靠DNA甲基转移酶(DNA methyltransferase,DNMT)来维持,哺乳动物细胞中目前发现维持甲基化的DNMT1、重新甲基化的 DNMT3A和 DNMT3B 3种 DNMT[14]。Zeb是DNMT抑制剂,可能主要通过抑制DNMT1,来诱导因甲基化失活的抑癌基因异常甲基化区域去甲基化而重新表达。在膀胱癌细胞中已证实Zeb能诱导p16去甲基化及重新表达[15]。
综上所述,本研究表明,启动子区异常甲基化是卵巢癌细胞RASSF1A基因失活的主要原因,去甲基化药物Zeb能逆转RASSF1A基因甲基化状态,调控RASSF1A基因重新表达,可能用于卵巢癌的临床治疗。
[1]AGATHANGGELOU A,COOPER W N,LATIF F.Role of the Ras-association domain family 1 tumor suppressor gene in human cancers[J].Cancer Res,2005,65(7):3497-3508.
[2]沈文静,戴冬秋,郭科军,等.上皮性卵巢癌组织中RASSF1A和BRCA1及p16基因异常甲基化检测及其临床意义的研究[J].中华肿瘤防治杂志,2008,15(7):530-533.
[3]MAKARLA P B,SABOORIAN M H,ASHFAQ R,et al.Promoter hypermethylation profile of ovarian epithelial neoplasms[J].Clin Cancer Res,2005,11(15):5365-5369.
[4]YOO C B,CHENG J C,JONESL P A.Zebularine:a new drug for epigenetic therapy[J].Biochemical Society Transactions,2004,32(6):910-912.
[5]CHENG J G,YOO C B,WEISENBERGER D J,et al.Preferential response of cancer cells to zebularine[J].Cancer Cell,2004,6(2):151-158.
[6]BAYLIN S B.Reversal of gene silencing as a therapeutic target for cancer-roles for DNA methylation and its interdigitation with chromatin[J].Novartis Found Symp,2004,259(1):226-233.
[7]沈文静,戴冬秋.卵巢癌表遗传学研究进展[J].中华肿瘤防治杂志,2007,14(10):790-794.
[8] WORM J,GULDBERG P.DNA methylation:an epigenetic pathway to cancer and a promising target for anticancer therapy[J].Oral Pathol Med,2002,31:443-449.
[9]沈文静,郭科军,戴冬秋,等.5-氮杂-2'-脱氧胞苷诱导卵巢癌细胞系p16基因去甲基化及表达增强[J].山西医药杂志,2007,36(9):781-784.
[10]APARICIO A,WEBER J S.Review of the clinical experience with 5-azacytidine and 5-aza-2'-deoxycytidine in solid tumors[J].Curr Opin Investig Drugs,2002,3:627-633.
[11]ORTIZ-VEGA S,KHOKHLATCHEV A,NEDWIDEK M,et al.The putative tumor suppressor RASSF1A homodimerizes and heterodimerizes with the Ras-GTP binding protein nore1[J].Oncogene,2002,21(6):1381-1390.
[12]姚晨,方娟娟,高丽丽,等.急性髓性白血病病人外周血中RIZ1的表达变化与启动子区甲基化相关性的研究[J].东南大学学报:医学版,2010,29(2):119-122.
[13]田小强,商延芳,黄培林.3种结肠癌细胞株中DLC-1基因的表达及其启动子区的甲基化状态研究[J].东南大学学报:医学版,2008,27(1):6-10.
[14]HERMAN J G,BAYLIN S B.Gene silencing in cancer in association with promoter hypermethylation[J].New Eng J Med,2003,349(8):2042-2054.
[15]CHENG J C,MATSEN C B,GONZALESF A,et al.Inhibition of DNA methylation and reactivation of silenced genes by zebularine[J].J Natl Cancer Inst,2003,95(5):399-409.
