西南喀斯特地区四种造林树种光合作用日变化及生态适应性研究
2011-04-16殷庆仓宋同清彭晚霞曾馥平
殷庆仓,宋同清,彭晚霞,曾馥平
(1.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,湖南长沙410125;2.广西大学林学院,广西南宁530004;3.中国科学院环江喀斯特生态系统观测研究站,广西环江547200)
西南喀斯特地区特殊的地理位置、气候条件、地质背景、水文结构及日益频繁的人类活动导致生态系统极易受损且难以恢复[1],受到人们广泛关注。为改善日益恶化的生态环境、提高人民的经济、文化、生活水平,同时配合好国家退耕还林还草政策的实施,西南喀斯特地区进行了长期、大量的生态恢复和重建工作。目前对生态系统退化机制、退化过程、生境条件、种群及更新特征[2,3,4]以及植被恢复中的土壤理化性质研究较多[5],而对树种的生态适应性研究相对缺乏。植被恢复是受损生态系统重建的一个重要环节,植被环境的耦合是森林植被恢复重建的最基本理论[6],森林生态系统的生态应当以构建各种具有高生物多样性、高功能、抗逆性强、稳定性强的森林生态系统类型为目标,保证系统能朝着良性的方向发展[7]。然而也有一些植被建设存在盲目性,缺乏森林生态恢复基础理论的科学指导,只注重短期的建设效果和显示度,与可持续发展的理念相违背。植物引种试验作为退化生态系统恢复和优化生态系统组建的重要内容,应得到足够重视。
植物光合作用是生态系统中最主要的同化过程,不仅在能量转换从而维持系统稳定、平衡大气碳氧中扮演了重要的角色,同时间接的反映了植物的生长状况。光合作用不仅受到植物种性的影响,也与环境因子息息相关,能比较直观地反馈植物对相关环境因子的适应信息。本文通过研究不同种植物的光合特性,为评价树种适生性、筛选优良树种和优化整治模式提供理论指导,以保证生态恢复中人工造林的科学性。
1 材料与方法
1.1 自然概况
试验在中国科学院环江喀斯特农业生态系统观测站植物引种园内进行,地理位置为 24°43′58.9″~24°44′48.8″N,108°18′56.9″~108°19′58.4″E,地势四周高,中间低,海拔最高为647.2m,最低为272.0m,坡度较陡,≥25°的坡面占62%。试验区为亚热带季风气候,年平均气温19.9℃,极端最高温度38.7℃,极端最低温度-5.2℃。年平均降雨量1389.1mm,降雨主要集中在4~8月,占全年降雨的73.7%,其他季节多干旱。年平均日照1451.1h,平均太阳总辐射量413.8kj/cm2,其中有效辐射量为211.9kj/cm2。植被覆盖率较低,多为喜钙耐旱的树种。
1.2 试验设计
供试材料为茶条木 (Delavaya toxocarpa)、樟叶槭(Acer albopurpurascens)、狗骨木(Cornus wilsoniana)、无患子(Sapindu smukorossi),分别属于无患子科、槭树科、山茱萸科、无患子科,均为资源引种植物,5年生,长势良好,生长在立地条件均一的坡脚上。每种树选取光照条件较好、生长状况相对一致的3株进行光合特性测定,每月测定一次光合速率日进程。
1.3 光合特性测定
2009年9月~2010年10月每月天气晴朗的时候,选择每株树向阳生长枝条上距枝顶第3到第5片完好成熟叶为待测叶片,在自然无风条件下利用美国Li-6400便携式光合测定系统于8:00~18:30时测定,每株树测定4片叶,每片叶重复测3次,每隔1小时测定一次,鉴于一次测量周期较长,为了尽量消除不同植物在一次测量中的光照差异,采用4种植物交叉测定的方法。测定内容包括净光合速率(Pn,μmolCO2/m2·s)、蒸腾速率(Tr,mmol H20/m2·s)、气孔导度(Gs,μmol H20/m2·s)、胞间 CO2浓度(Ci,μmolCO2/mol)、叶片温度(Tl,℃)、光合有效辐射强度(Par,μmol/m2·s)、大气 CO2浓度(Ca,μmolCO2/mol)、大气温度(Ta,℃)、大气相对湿度(Rh,%)等,并计算叶片的水分利用效率 (WUE,μmolCO2/mmol),即Pn/Tr。限于篇幅,本文选择天气状况稳定、光合作用较高的2010年8月份实验数据进行光合作用日变化分析。
1.4 数据处理
数据采用Excel-2003软件绘图、SPSS16.0软件进行统计分析。
2 结果与分析
2.1 不同树种净光合速率、蒸腾速率和水分利用效率日变化特征
植物的光合机构就像一台“生命的发动机”,而光合作用效率就是用来描述这台机器运转状况的指标和揭示光合机构运转调节规律的探针,也是植物生产力和作物产量高低的根本决定因素[8]。由表1和图1可见,不同树种的净光合速率(Pn)大小不同,顺序依次为茶条木>狗骨木>樟叶槭>无患子,前3种的差异不显著,均显著高于无患子,4种造林树种的Pn日变化呈相似的“双峰”曲线。曲线在8:00~10:00时迅速上升,10:00时出现第一个峰值,分别为7.31μmolCO2/m2·s、6.97μmolCO2/m2·s、5.69μmolCO2/m2·s和4.72μmolCO2/m2·s,然后急剧下降,均在11:00时出现低谷,“午休”现象明显,之后开始上升,茶条木和樟叶槭第二个峰值出现在12:00,狗骨木和无患子则出现在14:00,且较第一个峰值高,之后曲线逐渐下降,18:00时最低。相关研究表明:强光、高温、低湿和土壤干旱等环境条件引起气孔部分关闭和光暗呼吸加强或光合作用光抑制的发生导致了植物中午的光合“午休”现象[9,10]。此现象可以归结为气孔因素和非气孔因素两类,光合速率的降低可能是气孔导度降低的结果,也可能是气孔导度降低的原因,而只有当Pn和Ci同时减小可以证明光合速率的降低是气孔导度降低的结果[9],其余的光合速率降低要归因于叶肉细胞羧化能力的降低[11]。结合中午Pn、Ci的变化特征和Par、Ta、Rh实测值不难发现4种供试植物11:00光合速率降低并不是由于气孔导度降低引起的CO2供应不足所致,而是Par、Ta逐步升高导致叶肉细胞羧化能力下降的结果;12:00~15:00光合速率降低则主要是由于强光引起的气孔导度降低所致,与李新国等[12]的研究结果一致。

表1 不同植物净光合速率、蒸腾速率、水分利用效率日均值比较
蒸腾速率(Tr)的日变化曲线与Pn相似,但第二个峰值均明显高于第一个峰值,无患子、樟叶槭Tr的第二个峰值出现在13:00,茶条木和狗骨木出现在14:00,4个树种Tr的大小顺序为无患子、狗骨木、茶条木、樟叶槭,前三者的差异不显著,但均显著高于樟叶槭;蒸腾速率一天中最大值出现在13:00和14:00附近,此时Ta亦为一天中的最高值,Par也维持在较高水平,结合Tr与Ta、Par的相关分析结果,不难发现这是植物对强光、高温的一种适应性表现,温度通过调节叶面水汽压亏缺来调控蒸腾速率[13]。数据分析表明樟叶槭耐强光、高温的能力相对较强,而茶条木、狗骨木相对较弱,无患子较差。

图1 不同植物叶片净光合速率、蒸腾速率和水分利用效率日变化
叶片的水分利用效率可以用瞬时水分利用效率(WUE)和长期水分利用效率(植物吸收单位重量的水所产生的干物质的量)来表示。本次研究采用WUE,它往往反映了一定条件下植物碳同化和水分的平衡关系[14,15]、适应性强弱等[16,17,18]。如图1所示,4种造林树种的水分利用率(WUE)总体上均呈下降趋势,表明上午光合速率的增加幅度小于蒸腾速率的增加幅度,而下午光合速率的降低幅度大于蒸腾速率降低的幅度;茶条木和狗骨木在17:00时出现了轻微的回升现象,同此时大气CO2升高相似,与Curtis等[19]的研究相一致,表明大气CO2升高在一定程度上促进了水分利用效率的提高。WUE的顺序为茶条木>樟叶槭>狗骨木>无患子,前3者差异不显著,均显著高于无患子。
2.2 光合生态因子日变化
由图2可知,Par、Ta的日变化剧烈,上午随时间的推移急剧上升,12:00时最高,峰值为1064.67μmol/m2·s,然后急剧下降,18:00时为41.82μmol/m2·s;Ta的日变化曲线平缓,且随Par的增加而升高,但明显的滞后于辐射,峰值出现在14:00,为 33.95 ℃;Ca、Rh 与 Par、Ta正好相反,日变化为不规则的“V”字型曲线,8:00时由于Par低、光合结构活化较慢,光合碳同化较弱,前一晚植物呼吸释放了大量的CO2,Ca值最高,为 391.98 μmolCO2/mol,随后急剧下降,11:00~16:00时徘徊在较低水平,最低值出现在16:00,为375.49 μmolCO2/mol,Rh与Ta呈强烈负相关,最低值出现在14:00,为33.75mbar。
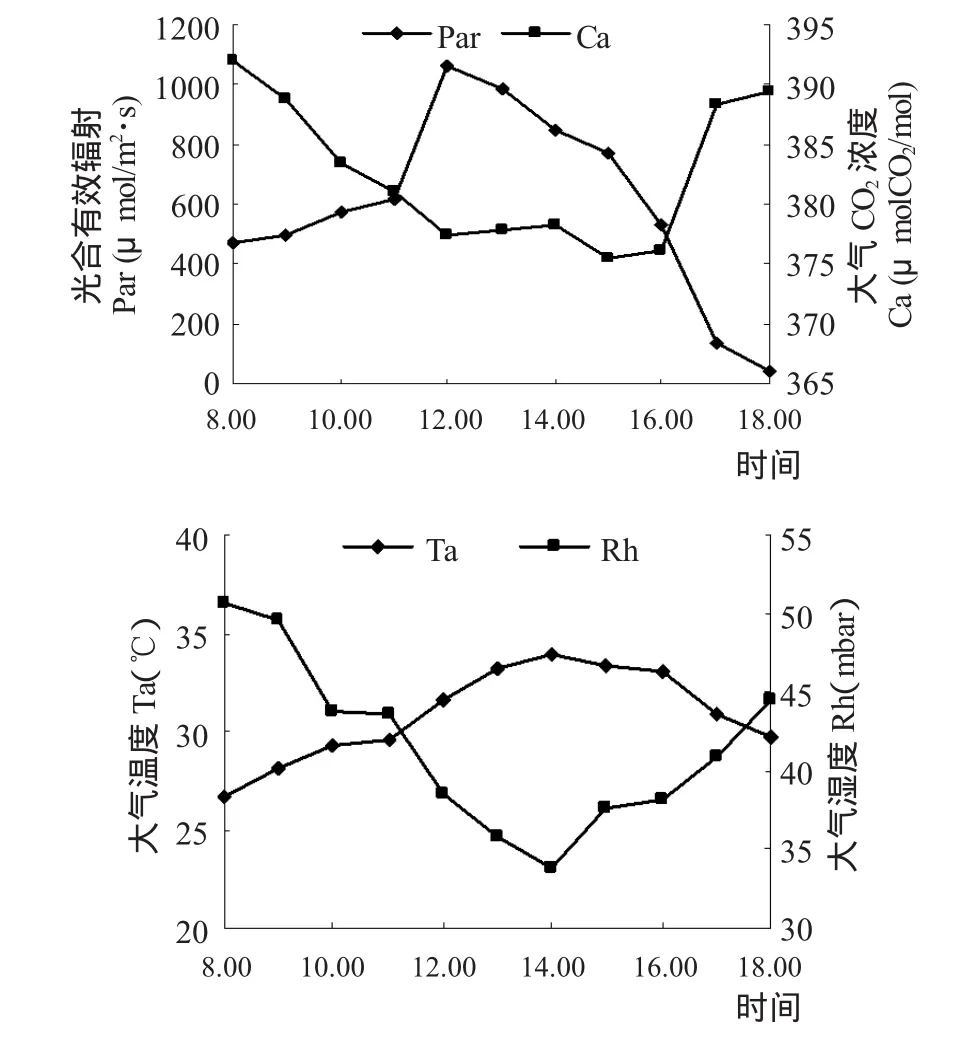
图2 植物生长环境因子日变化
2.3 气孔导度和胞间CO2浓度日变化特征
Gs是衡量叶片内外CO2扩散阻力大小的指标,控制着叶片内外CO2浓度[11]。如图3所示,4种植物的Gs和Ci日均值大小顺序为无患子>狗骨木>茶条木>樟叶槭,与Tr日均值一致。Gs日变化呈不明显的高—低—次高的“双峰”曲线,第一峰值均出现在11:00,无患子和狗骨木的低谷在12:00,第二峰值在13:00,茶条木和樟叶槭的低谷在13:00,第二峰值出现在14:00。有关Gs变化规律现有多种说法:早晨和傍晚低Par导致Gs在早晨和傍晚较低[20];Gs随着Par和Ta的增加而增加[21];对Gs影响最大的环境因子是Rh[22]。结合Par、Ta、Rh和Gs的变化特征发现,4种植物Gs呈规律性变化是Rh同Par、Ta综合作用的结果,至少是受Rh与Par、Ta其中之一共同影响的。受Gs变化的影响,Ci值8:00后均下降,11:00有所回升后又下降,14:00出现最低值后开始回升,18:00到达较大值。
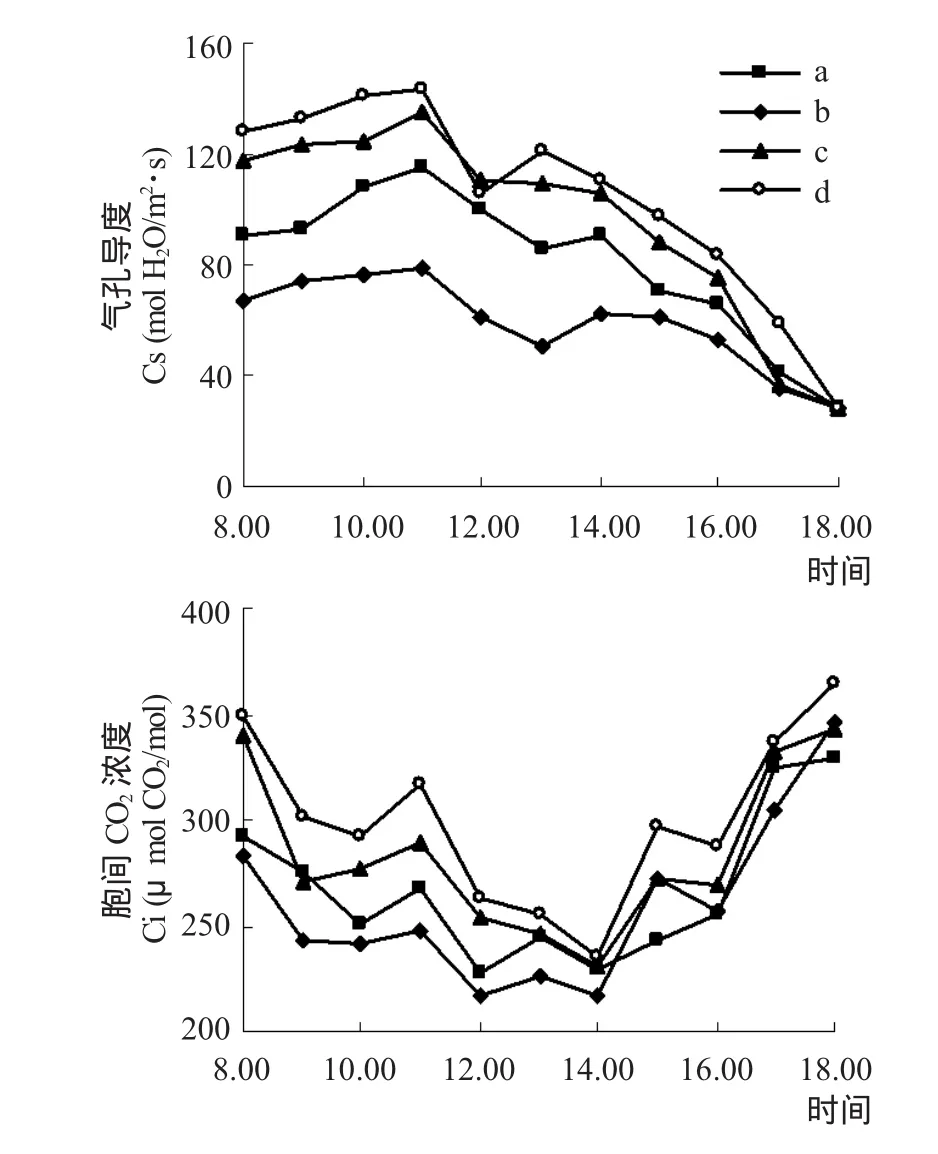
图3 不同植物叶片气孔导度和胞间CO2浓度日变化
2.4 光合速率、蒸腾速率与生态生理因子相关性分析
由表2可知,4种植物的Pn均与Par、Gs呈极显著正相关,与Ci呈极显著负相关,与Ta、Rh的相关性不大;Tr与Par、Ta、Gs呈正相关,与Par相关性极显著,除樟叶槭外,Tr与Gs相关性均达到显著或极显著,与Ta的相关性不显著。Tr与Ci、Rh之间呈负相关,与Ci相关极显著,与Rh的关系只有无患子达到了显著水平。表明除樟叶槭外对Pn、Tr影响最强的生理生态因子为Par、Gs和Ci,而Ta、Rh对其影响相对较小。

表2 四种植物的净光合速率、蒸腾速率与生理生态因子的相关系数
3 结论与讨论
3.1 结论
4种造林树种的Pn日变化呈相似的“双峰”曲线。中午强光、高温、低湿度引起的叶肉细胞羧化能力下降均在不同程度上导致了Pn的降低,11:00时均出现了光合“午休”现象,这是植物在长期进化过程中形成的一种自我保护方式,以减少光合机构的光破坏;12:00~15:00时Pn下降则主要为强光引起的Gs降低所致。此外,4种供试植物的Tr日变化趋势亦为“双峰”曲线。温度通过调节叶面水汽压亏缺来调控蒸腾速率,在Ta最高时Tr达到了最大值;WUE则在整个测定周期中大体呈递减趋势,而下午晚些时候大气CO2浓度升高在一定程度上促进了狗骨木和茶条木水分利用效率的提高。
数据分析表明:对Pn影响最为显著的生态生理因子为Par、Gs,其次为 Ci、Rh、Ta;茶条木和狗骨木对强光、高温、干旱适应性相对较强,其次为樟叶槭和无患子。综合考虑西南喀斯特地区特殊的地质地貌、气候、水文条件等,可认为茶条木、狗骨木是具有较高光合速率、水分利用效率的植物,能较好的与该地区的生态环境相适应,樟叶槭其次,而无患子适宜度最差,属于耗水型树种。
3.2 讨论
西南喀斯特地区特殊的气候、地质背景,使该区的水、热、光时空分布极其不均匀。虽然雨热同期,但旱、雨季区分明显,夏季时晴时雨,天气变化频繁,常常导致局部高温和干旱,水、热、光因此也成了限制植物生长的重要因素。故耐干旱、耐高温、耐强光的植物将更有利于生存,适应性特征表现为具有高水分利用效率、低蒸腾速率和高光合速率。具有这些特征的植物既能很好的抵御干旱胁迫,又能最大限度的利用太阳光能,同时避免了光合机构的光损伤,最大限度的利用太阳光能同化CO2并充分利用有限的水分,有利于植被群落朝着稳定性强、抗逆性强的方向发展,与可持续的生态恢复理念相符合,可考虑在该地区种植。而低水分利用效率、高蒸腾速率、低光合速率的植物适生性相对较差,在夏季容易受到强光、高温等胁迫,在水分利用方面处于劣势,用作造林树种应慎重。建议采取套种等复合种植模式来改善其生境小气候,或种植在相对背光的坡脚,以提高造林植物的存活率。本文基于对4种造林树种叶片气孔气体交换相关特征和环境因子日变化的研究,对4种植物的生态适应性做了初步评价,其适应性顺序为:茶条木>狗骨木>樟叶槭>无患子。
[1] 李兵阳,王世杰.喀斯特石漠化的研究现状与存在的问题[J].地球环境,2006,34(3):9-14.
[2]曾馥平.西南喀斯特脆弱生态系统退化原因与生态重建途径[J].农业现代化研究,2008,29(6):672-675.
[3] 任海.喀斯特山地生态系统石漠化过程及其恢复研究综述[J].热带地理,2005,25(3):195-200.
[4] 喻理飞,朱守谦,叶镜中,等.人为干扰与喀斯特森林群落退化及评价研究[J].应用生态学报,2002,13(5):529-532.
[5] 何永彬,张信宝,文安邦.西南喀斯特山地的土壤侵蚀研究探讨[J].生态环境学报,2009,18(6):2393-2398
[6] 赵文智,程国栋.干旱区生态学水文过程研究若干问题评述[J].科学通报,2001,46(22):1851-1857.
[7] 赵平.退化生态系统植被恢复的生理生态学研究进展[J].应用生态学报,2003,14(11):2031-2036.
[8] 许大全.光合作用效率[M],上海:上海科学技术出版社,2002,204.
[9] 许大全.光合作用测定及研究中一些值得注意的问题[J].植物生理学通讯,2006,42(6):1163-1167.
[10]许大全,张玉忠,张荣铣.植物光合作用的光抑制[J].植物生理学通讯,1992,28(4):237-243.
[11]FarqubarGD,SharkeyTD.Stomstalconductanceand photosynthesis[J].Ann Bey PlantPhysiol,1982(33):317-345.
[12]李新国,许大全,孟庆伟.银杏叶片光合作用对强光的响应[J].植物生理学报,1998,24(4):354-360.
[13]夏尚光,张金池,梁淑英.南方岩榆光合作用日变化及其影响因子研究[J].亚热带科学,2007,36(3):8-11.
[14]孙谷畴,赵平,蔡锡安,等.马占相思叶片液汁碳同位素甄别率和水分利用效率[J].生态学杂志,2008,27(4):497-503.
[15]苏培玺,张立新,杜明武,等.胡杨不同叶形光合特性、水分利用效率及其对加富CO2的响应[J].植物生态学报,2003,27(1):34-40.
[16]迟丽华,宋凤斌.松嫩平原4种植物光合作用光响应特性研究[J].吉林农业大学学报,2007,29(2):119-122.
[17]刘金祥,麦嘉玲.CO2浓度增强对沿阶草光合特性的影响[J].中国草地,2004,26(3):l3-18.
[18]杨建伟,梁宗锁,韩蕊莲,等.不同干旱土壤条件下杨树的耗水规律及水分利用效率研究[J].植物生态学报,2004,28(5):630-636.
[19]CurtisPS,WangXZ.Ameta-analysisofelevated CO2effectsonwoody plantmass,form and physiology[J].Oecologia,1998(113):299-313.
[20]王玉辉,周广胜.松嫩平原盐碱化草地羊草叶片生理生态特性分析[J].应用生态学报,2000,11(3):12-19.
[21]温达志,周国逸,张德强,等.四种禾本科牧草植物蒸腾速率与水分利用效率的比较[J].热带亚热带植物学报,2000(增刊):67-76.
[22]项斌,林舜华,高雷明.紫花苜蓿对CO2倍增的反应:生态生理研究和模型拟合[J].植物学报,1996,38(1):63-71.
