纤维素结合域的酿酒酵母表面展示及其黏附位点初探
2011-03-14张玉杰王佳堃叶均安刘建新
张玉杰 王佳堃 叶均安 刘建新
(浙江大学奶业科学研究所,杭州 310029)
反刍动物通过瘤胃微生物发酵纤维素、半纤维素等结构性碳水化合物满足机体的能量需要。瘤胃微生物对纤维物质的黏附是降解纤维的首要条件[1-2]。因此,微生物对纤维物质的黏附一直是反刍动物营养研究的热点之一。微生物对纤维物质的黏附包括非特异性黏附和特异性黏附 2种形式,特异性黏附又以酶的纤维素结合域(cellulosebinding dom ain,CBD)研究得相对深入[3]。CBD是碳水化合物活性酶内具有纤维素结合能力的高度折叠的一段氨基酸序列,目前的研究致力于CBD的应用[4-7]、蛋白质和碳水化合物结合机制[8]等,较少涉及瘤胃优势纤维分解细菌。产琥珀酸丝状杆菌(Fibrobacter succinogenes)是瘤胃内主要的三大纤维分解细菌之一[9-11]。CAzy(http://www.cazy.org)数据库显示,产琥珀酸丝状杆菌 S85的 CBD分属于家族 2、6、11和 30,而这 4个家族在微生物中的分布并不均衡,很多细菌酶都具有 CBD2结构域,突显了 CBD2的重要性。CBD是高度折叠的氨基酸序列,传统的研究方法难以应用于其研究。酵母表面展示技术是近年来发展较快的一种真核蛋白表达系统,其基本原理是将外源靶蛋白基因与特定的载体基因序列融合后导入酵母细胞,利用酵母细胞内蛋白转运系统转运到膜表面,从而使靶蛋白固定化表达在酵母细胞表面[12-13]。酿酒酵母对表达蛋白翻译后修饰加工,易于培养、传代稳定[13],在研究 CBD的过程中无需分离、纯化操作,而且表达的蛋白可通过特异性免疫组化进行灵敏而方便地检测和筛选。为此,本试验利用酿酒酵母展示技术展示产琥珀酸丝状杆菌的 CBD2,并通过特异性免疫组化反应分析 CBD 2纤维物质的黏附位点,探讨其重要性,为后续深入研究 CBD与瘤胃细菌特异性纤维黏附位点的关系研究奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒、外源靶蛋白基因和植物材料
酿酒酵母 EBY 100和质粒 pYD 1均由杭州师范大学陈振明副教授馈赠。Trans5α感受态细胞购于北京全式金生物技术有限公司。产琥珀酸丝状杆菌 S85的 CBD2与 pUC57质粒的合成由上海捷瑞生物工程有限公司完成。稻草取自浙江大学试验牧场。
1.1.2 酶与试剂
限制性内切酶 Bam HⅠ和 XhoⅠ及 T4 DNA连接酶购于 Fermentas公司。核糖核酸酶 A(RNase A)、核酸分子质量标准(500~15 000和100 bp)和割胶回收试剂盒购自于 TaKaRa公司。二甲基亚砜(DMSO)、聚乙二醇 -3350(PEG-3350)、鲑鱼精 DNA购于 Sigma公司。抗 V 5异硫氰酸荧光素标记抗体(Anti-V 5-FITC)购于 Invitrogen公司。其它生化试剂为国产分析纯级。
1.1.3 培养基和缓冲液
LB培养基:蛋白胨 10 g/L、酵母膏 5 g/L、NaCl10 g/L、琼脂粉 15 g/L,用于 Trans5α培养。
LB氨苄青霉素固体选择性培养基:LB培养基中加入氨苄青霉素(ampicillin)100 mg/L、琼脂粉 15 g/L,用于重组质粒的筛选。
酵母膏胨葡萄糖(YPD)培养基:蛋白胨20 g/L、酵母膏 10 g/L、葡萄糖 20 g/L、琼脂粉15 g/L,用于酿酒酵母 EBY 100培养。
无氨基酸酵母氮源 -酸水解酪素(YNBCAA)培养基 :YNB 6.7 g/L、CAA 5 g/L、葡萄糖20 g/L、琼脂粉 15 g/L,用于转化子的转接或诱导。
MD转化培养基:YNB 6.7 g/L、亮氨酸0.1 g/L、葡萄糖 20 g/L、琼脂粉 15 g/L,用于酵母转化子的筛选。
PEM缓冲液:哌嗪 -N,N′-双(2-乙磺酸)(PIPES)50 mmol/L、乙二胺四乙酸 (EDTA)5 mm ol/L、M gSO45 mmo l/L,pH 6.9,用于稻草片固定。
磷酸盐缓冲液(10mm ol/L):NaCl 8 g/L、KCl 0.2 g/L、Na2HPO41.44 g/L、KH2PO40.24 g/L,pH 7.4,用于稻草片的制备和孵育等。
1.2 重组质粒 pMCBD2的构建
在 LB氨苄青霉素固体选择性培养基上划线培养,获得含 CBD2基因的大肠杆菌 Trans5α单菌落。挑取单菌落 37℃,200 r/min过夜扩增,十二烷基硫酸钠(SDS)碱裂解法抽提质粒[14],Bam HⅠ和 XhoⅠ双酶切质粒,电泳后割胶回收获得CBD2基因序列。用同样的方法获得开环 pYD1序列。将 2段序列经 T4 DNA连接酶连接后,连接产物转化大肠杆菌 Trans5α,活化后涂布于 LB氨苄青霉素固体选择性培养基。挑取筛选出的单菌落经液体选择性培养基扩增后,抽提质粒,通过限制性酶切分析和测序(上海桑尼公司)验证重组质粒构建是否成功。
1.3 酵母转化及诱导表达
采用醋酸锂法[15]将构建成功的质粒 pMCBD2转化酿酒酵母细胞 EBY 100,转化后涂布于 MD培养基。挑取筛选出的单菌落,接种于 10 m L YNBCAA培养基中,30℃、200 r/min培养过夜。待菌体生长到 OD600nm在 2~5时,5 000×g离心 5m in收集菌体。用 2%半乳糖的 YNB-CAA培养基重新悬浮菌体,使菌液 OD600nm为 0.5~1.0,20℃条件下 200 r/min摇床培养 6 h诱导基因表达。
1.4 稻草片的制备与酵母细胞孵育
改进 Wang等[16]和 McCartney等[17]的方法制备植物材料,并对其进行酵母细胞孵育。具体做法如下:取整株稻草,剥离茎、叶片和叶鞘,在茎的第二节上缘,节下 2厘米处截取长为 3厘米的茎片断,纵切为 6份,水浸过夜后切割 100~300μm的横切片断。在含 4%多聚甲醛的 PEM缓冲液中固定 1 h后,用 10 mm ol/L磷酸盐缓冲液冲洗。取1.5m L诱导表达的重组酵母细胞菌液,5 000×g离心 5m in获得重组酵母细胞,用 10mmol/L磷酸盐缓冲液洗涤细胞后,加入 250μL上述磷酸盐缓冲液制得细胞悬液,与稻草片一起室温条件下孵育1.5 h。
1.5 免疫组化反应
孵育后的稻草片用 10 mmol/L磷酸盐缓冲液清洗 3次,以除去非特异黏附的酵母细胞。随后用 1∶300稀释的抗 V5异硫氰酸荧光素标记抗体处理稻草片 1.5 h,再用 10 mmol/L磷酸盐缓冲液清洗 3次后,荧光显微镜(尼康 ECLIPSS 80i)下检测。
2 结 果
2.1 重组质粒 pMCBD2和重组菌株 pMCBD 2-EBY 100的构建
pYD1是 5 009 bp的环状 DNA,CBD2长为322 bp通过 pUC57携带。利用 Bam H I和 Xho I将目标 CBD2序列克隆至表面展示载体 pYD1的AGA 2 3′端,使 AGA 2-CBD 2融合基因处于 GAL1启动子下游,构建载体 pMCBD2,以实现半乳糖诱导下的高效表达(图 1A)。
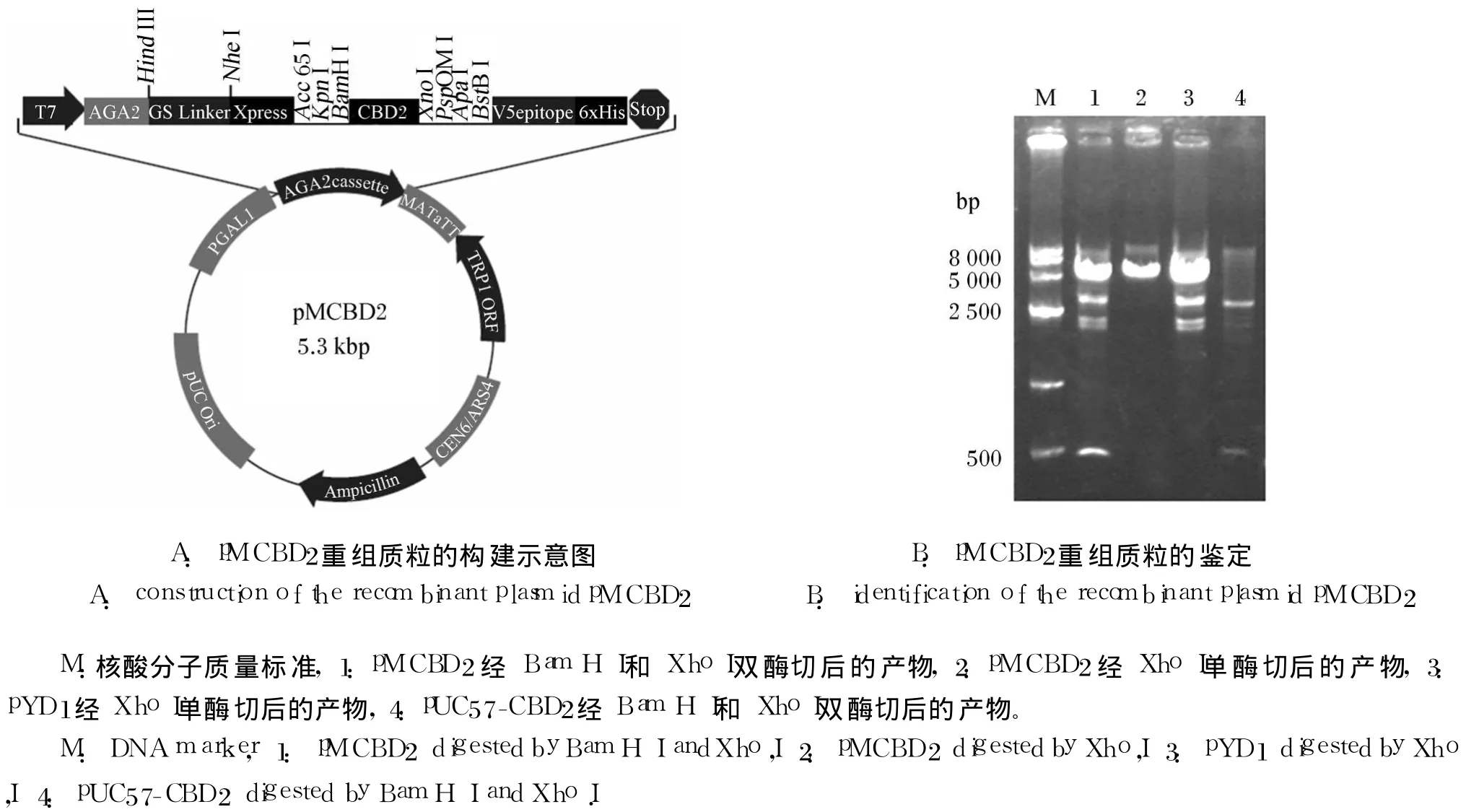
图 1 pM CBD 2重组质粒的构建及鉴定Fig.1 Construction and identification of the recombinant p lasm id pMCBD 2
pMCBD 2和 pYD 1双酶切图谱均出现多个条带,表明酶切不完全,但 2者比对可看到 pMCBD2携带了约 500 bp的片段(图 1B)。
pMCBD 2单酶切仅获得开环序列,测序分析的结果(图 2)进一步证实重组质粒构建成功。
2.2 重组菌株 pMCBD2-EBY 100纤维黏附位点分析
因为 pYD1含有 V 5抗原表位,所以重组的pMCBD2-EBY 100酵母细胞在诱导表达后,可借助免疫组化,通过荧光监测 CBD 2的纤维黏附位点。免疫组化结果显示,未重组 CBD 2的 pYD 1-EBY 100不具有黏附稻草的能力(图 3A),而重组的 pMCBD2-EBY 100对稻草的厚壁组织、薄壁组织和维管组织均有黏附。黏附程度随稻草组织结构的变化而不同,细胞相对致密的厚壁和维管组织的荧光强度大于细胞相对疏松的薄壁组织(图 3B)。
3 讨 论
酿酒酵母细胞壁蛋白 a凝集素有 2个亚基,其中 1个是 AGa2,AGa2-CBD2融合蛋白通过二硫键与酿酒酵母细胞壁蛋白 a凝集素的另一亚基AGa1蛋白结合,然后 AGa1蛋白通过糖基磷脂酰肌醇(GPI)共价结合到细胞壁上。因为酿酒酵母EBY 100基因组中被整合了启动子 gall-AGal开放阅读框,所以 AGa2-CBD2融合蛋白能与充足的AGa1蛋白结合,使 CBD2在酿酒酵母细胞表面展示。
稻草的细胞壁在细胞发育过程中,由于原生质体代谢活动不断改变,所形成的壁物质存在组成种类、数量、比例以及物理性质的差异,使其在显微镜下呈现出自外而内的胞间层、初生壁和次生壁的 3层结构[18]。纤维素约占稻草细胞壁的60%,分布于初生壁和次生壁,且主要以微纤维组成的结晶形状存在。半纤维素则主要是以木聚糖和葡萄糖醛酸的缩合物形式存在,且随初生壁向次生壁木质化程度减弱而减少[19]。基于结构和功能差异,CBD2可划分为 A和 B 2种类型。A型为黏附结晶纤维素,B型为黏附多聚糖链[8]。图 4序列比对及图 5利用 SW ISS-MODEL/SW ISS-Pdb-View er[20-22]预测的蛋白质三级结构结果显示产琥珀酸丝状杆菌的 CBD2具有 3个暴露于空间结构外侧的色氨酸(图 4中黑框标出的“W”),且与第1个色氨酸相隔 2个位置的氨基酸为甘氨酸(图 4中“*”标出的“G”)。依据这一结果理论推测产琥珀酸丝状杆菌的 CBD2可能属于类型 A[23],即该 CBD特异性地黏附结晶纤维素,而对木聚糖无黏附能力。这一推测与显微镜下观察到产琥珀酸丝状杆菌的 CBD2对厚壁、薄壁和维管组织细胞均有黏附,且荧光强度与细胞壁的厚度一致相吻合(图 3)。Blake等[24]以烟草茎为底物证实CBD2A细胞壁黏附范围和强度高于 CBD1、CBD 3A、CBD10等,尤其是 CBD 2A可黏附木质部的次生壁。上述结果表明利用酵母表面展示系统和免疫组化分析技术可研究瘤胃微生物 CBD的黏附特性。

图 2 pM CBD 2与 CBD 2序列比对结果Fig.2 A comparison betw een sequences of pMCBD 2 and CBD 2
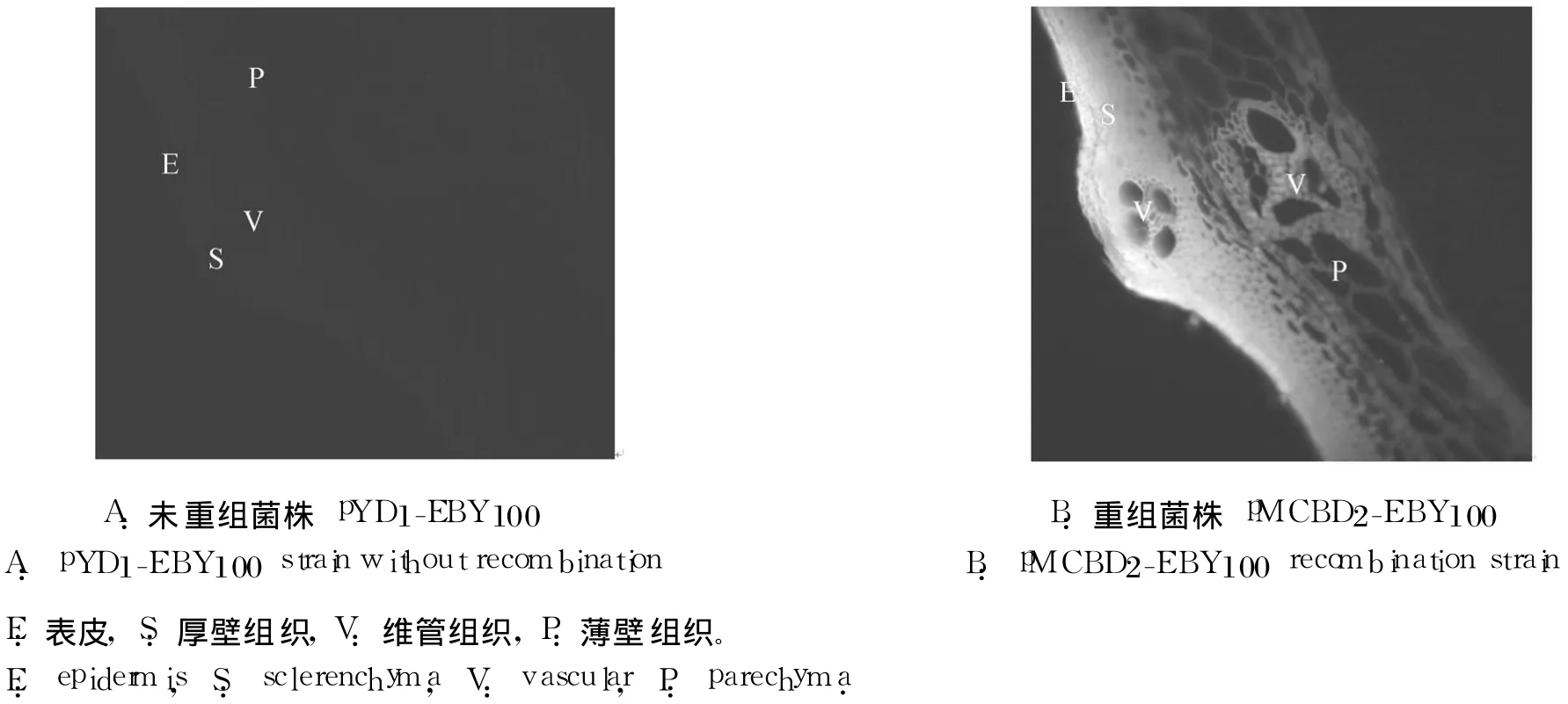
图 3 免疫组化观测 CBD 2在稻草茎横切面上的黏附位点Fig.3 Immunohistochem isty detection of CBD 2binding locus on transverse sections o f straw stem

图 4 CBD 2A多序列比对Fig.4 Sequence alignment of CBD 2A

图 5 预测的产琥珀酸丝状杆菌 CBD 2三维结构Fig.5 A p rediction o f three dimensional structure of CBD 2 of F.succinogenes
4 结 论
本试验利用酿酒酵母表面展示系统成功表达了产琥珀酸丝状杆菌 CBD2,免疫组化分析显示产琥珀酸丝状杆菌 CBD2的黏附位点广泛,可黏附薄壁、厚壁和维管组织。酿酒酵母表面展示技术与免疫组化技术联用研究瘤胃细菌 CBD的黏附位点是可行的。
[1] GONG J,FORSBERG CW.Factors affecting adhesion Fibrobacter succinogenes S85 and adherence defectivemutant to cellulose[J].Applied and EnvironmentalM icrobiology,1989,55:3039-3044.
[2] FORSBERG CW,FORANO E,CHESSON A.M icrobial adherence to the plant cell w all and enzymatic hydrolysis[C]//CRONJE P D.Rum inant physiolo-gy:digestion,metabolism,grow th and reproduction.Proceeding of the 9th ISRP.UK:CAB International,2000,79-97.
[3] MIRON J,BEN-GHEDALIA D,MORRISON M.Invited review:adhesion mechanisms of rumen cellulolytic bacteria[J].Journal of Dairy Science,2001,84:1294-1309.
[4] FUJITA Y,KATAH IRA S,UEDA M,et al.Construction o f whole-cell biocatalyst for xylan degradation through cell-surface xylanase display in Saccharomyces cerevisiae[J].Journal of Mo lecular Catalysis B:Enzymatic,2002,17:189-195.
[5] FUKUDA T,ISHIKAW A T,OGAWA M,et al.Enhancement o f cellulase activity by clones selected from the combinatorial library of the cellulose-binding dom ain by cell surface engineering[J].Biotechnology Progress,2006,22:933-938.
[6] MURAI T,UEDA M,KAWAGUCHI T,et al.Assim ilation of celloo ligosaccharides by a cell surfaceengineered yeast expressingβ-glucosidase and xarboxymethy lcellulase from Aspergillus aculeatus[J].App lied and Environmental M icrobiology,1998,64:4857-4861.
[7] NAM JM,FUJITA Y,ARAIT,et al.Construction of engineered yeastw ith theability of binding to cellulose[J].Journal of M o lecu lar Catalysis B:Enzymatic,2002,17:197-202.
[8] BORASTON A B,BOLAM D N,GILBERT H J,et al.Carbohyd rate-binding modules:fine-tuning polysaccharide recognition[J].Biochem ical Journal,2004,382:769-781.
[9] FORSBERG CW,CHENG K J,WHITE BA.Polysaccharide degradation in the rumen and large intestine[M]//MACK IE R I,BRUCE A.WHITE R E,et al.Isaacson gastrointestinal m icrobiology.New York:Chapman and Hall,1997,319-379.
[10] LATHAMM J,BROOKER B E,PETIPHER G L,et al.Ruminococcus flavefaciens cell coat and adhesion to cotton cellulose and cell walls in leaves o f perennial ryegrass[J].App lied and Environmental M icrobiology,1978,35:156-165.
[11] DINSDALE D,MORRISE J,BACON JSD.Electron m icroscopy of the m icrobial populations present and their modes of attack on various cellu losic substrates undergoing digestion in the sheep rumen[J].App lied and Environmental M icrobio logy,1978,36:160-168.
[12] KONDO A,UEDA M.Yeast cell-surface display-applications ofmolecular display[J].Applied M icrobiology and Biotechnology,2004,64:28-40.
[13] SH IBASAK I S,MAEDA H,UEDA M.Molecular disp lay technology using yeast-arm ing technology[J].Analytical Sciences January,2009,25:41-49.
[14] SAMBROOK J,RUSSELL D W.分子克隆实验指南[M].3版.黄培堂,王栋樑,周晓巍,等,译.北京:科学出版社,2002.
[15] ITOH,FUKUDA Y,MURATA K,et al.Transformation of intact yeast cells treated with alkali cations[J].Journal of Bacteriology,1983,153(1):163-168.
[16] WANG J K,LIU J X,LI J Y,et al.Histological and rumen degradation changes of rice straw stem epiderm is as influenced by chem ical pretreatment[J].Animal Feed Science and Technology,2007,136:51-62.
[17] MCCARTNEY L,BLAKE AW,FLINT JE,et al.D ifferential recognition o f plant cell walls bym icrobial xy lan-specific carbohydrate-bindingmodules[J].Proceedings o f the National A cademy of Sciences o f the United States of America,2006,103(2):4765-4770.
[18] WANG JK,CHEN X L,LIU JX,et al.Histological changes of tissues and cellw all of rice straw influenced by chem ical pretreatments[J].Asian-Australasian Journal o f Animal Sciences,2008,21:824-830.
[19] 王佳堃,叶丹妮,李文婷,等.化学预处理对稻草硅化程度的影响及其与稻草降解率的关系研究[J].动物营养学报,2008,20(2):170-175.
[20] ARNOLD K,BORDOLI L,KOPP J,et al.The SW ISS-MODEL w orkspace:a w eb-based environment for p rotein structure homo logy modeling[J].Bioinformatics,2006,22:195-201.
[21] SCHWEDE T,KOPP J,GUEX N,PEITSCH M C.SW ISS-MODEL:an automated p rotein homologymodeling server[J].Nucleic Acids Research,2003,31:3381-3385.
[22] GUEX N,PEITSCH M C.SWISS-MODEL and the Swiss-PdbViewer:an environment for com parative protein modeling[J].Electrophoresis,1997,18:2714-2723.
[23] SIMPSON P J,XIE H,BOLAM D N,et al.The structural basis for the ligand specificity of fam ily 2 carbohyd rate-bindingm odules[J].The Journal of Biological Chem istry,2000,275:41137-41142.
[24] BLAKE A W,MCCARTNEY L,FLINT JE,et al.Understanding the bio logical rationale for the diversity of cellulose-directed carbohyd rate-binding modules in prokaryotic enzym es[J].The Journal of Biological Chem istry,2006,281:29321-29329.
