饲粮添加不同碳链长度脂肪酸对泌乳奶牛生产性能和乳脂肪酸组成的影响
2011-03-14王加启李发弟卜登攀赵小伟徐晓燕周凌云
崔 海 王加启* 李发弟 卜登攀 赵小伟 徐晓燕 孙 妍 周凌云
(1.甘肃农业大学动物科学技术学院,兰州 730070;2.中国农业科学院北京畜牧兽医研究所,动物营养学国家重点实验室,北京 100193)
研究表明,外源添加富含长链不饱和脂肪酸的油脂或脂肪酸可显著提高奶牛乳脂中不饱和脂肪酸含量,但对乳脂率没有提高,甚至会出现乳脂率降低的情况[1]。在这种情况下,乳脂中短链和中链脂肪酸(short-and medium-chain fatty acids,SMCFA)含量降低的幅度显著高于长链脂肪酸(long-chain fatty acids,LCFA)[2]。反刍动物乳脂中脂肪酸有2个来源,4~14碳脂肪酸及约50%的16碳脂肪酸(即本文中所述的SMCFA)在乳腺中通过从头合成途径合成,约50%的16碳脂肪酸和大于16碳的脂肪酸(即本文中所述的LCFA)则来源于血脂[3]。研究表明,十二指肠灌注含有SMCFA的黄油使乳脂率显著高于对照组,并且认为SMCFA是提高乳脂率的重要因素[4]。本研究拟以荷斯坦奶牛典型乳脂脂肪酸组成为参考,模拟SMCFA和LCFA的脂肪酸组成,并且在饲喂试验中添加量保持一致,以对不同碳链长度脂肪酸在奶牛乳脂合成中的贡献进行比较。
1 材料与方法
1.1 试验动物与试验设计
选择北京沧达福牛场36头泌乳中期[(183±46)d]、胎次和泌乳量相近、健康状况良好的中国荷斯坦奶牛,采完全随机区组试验设计,将奶牛分为对照组、SMCFA组和LCFA组,每组12个重复,每个重复1头牛,均饲喂相同的基础饲粮,其中SMCFA组和LCFA组奶牛分别补饲400 g/d SMCFA和400 g/d LCFA。
1.2 饲粮组成及饲养管理
试验用基础饲粮根据NY/T 34—2004奶牛营养需要配制,精粗比为50∶50,其组成及营养水平见表1。SMCFA和LCFA组成分别模拟荷斯坦奶牛典型乳脂中SMCFA和LCFA组成配制。因中国奶牛品种绝大部分为荷斯坦奶牛,且黄油是从鲜牛乳中提炼出的成,所以黄油中脂肪酸组成可代表荷斯坦奶牛典型乳脂脂肪酸组成,因此将黄油中脂肪酸组成作为参考值模拟SMCFA和LCFA中脂肪酸比例[5]。SMCFA、LCFA和黄油的脂肪酸组成见表2。
试验牛经过7 d预试期后转入试验阶段,正试期为56 d,试验牛采用拴系式饲养,饲粮分3次饲喂(08:30、14:30和20:30),试验牛每天挤奶3次(08:00、14:00 和20:00)。
1.3 样品处理与指标测定
采样方法:试验前以及正试期每周各采集1次乳样,早、中、晚样按照4∶3∶3的比例混合制样。将混合后的乳样分为2部分,一部分乳样添加0.05%布罗波尔防腐剂于4℃保存用于分析乳成分;另一部分乳样于-20℃保存用于分析乳脂脂肪酸组成。
检测指标:乳成分在农业部奶及奶制品质量监督检验测试中心通过M ilk Scan FT120 Foss electric型乳样自动分析仪检测。乳脂脂肪酸组成采用气相色谱法(HP6890,SP-2560毛细管柱:100 m ×0.25 mm,内径 0.2 μm,FID 检测器),以C17∶0为内标,使用二阶程序升温法分离检测,详细分析方法参见文献[6]。
1.4 数据分析
试验数据经 Excel初步处理后,采用 SAS V8.2软件进行统计分析,并采用PROC M IXED进行方差分析,以试验牛泌乳日龄为协变量,数据为最小二乘均值,显著性水平为 P<0.05,采用Tukey法进行多重比较。
2 结果
2.1 饲粮添加不同碳链长度脂肪酸对泌乳奶牛生产性能的影响
由表3可知,试验前各组乳产量、乳成分含量及产量均差异不显著(P>0.05)。由表4可知,饲粮添加不同碳链长度脂肪酸后,SMCFA组乳脂率显著高于 LCFA 组(P <0.05),提高 14.62%;而对照组乳脂率与其他2个组差异均不显著(P>0.05)。乳产量、乳蛋白率、乳糖率、乳总固形物含量和乳成分产量在3个组间均差异不显著(P>0.05)。

表1 基础饲粮组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of the basal diet(DM basis) %

表2 SMCFA、LCFA和黄油的脂肪酸组成Table 2 Fatty acid composition of SMCFA,LCFA and butter %
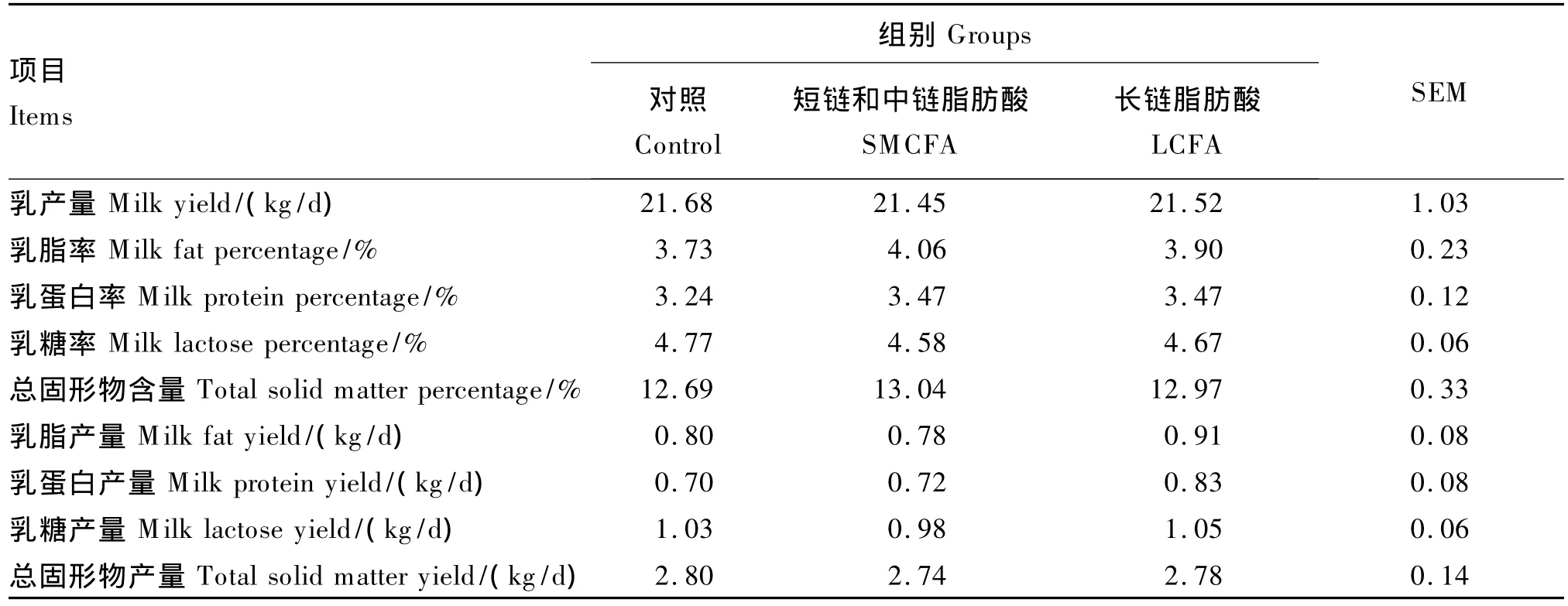
表3 试验前各组泌乳奶牛的生产性能Table 3 M ilk yield of lactating dairy cows in each group before experiment

表4 饲粮添加不同碳链长度脂肪酸对泌乳奶牛生产性能的影响Table 4 Effects of supplemental fatty acids w ith different carbon chain lengths on m ilk yield of lactating dairy cows
2.2 饲粮添加不同碳链长度脂肪酸对泌乳奶牛乳脂肪酸组成的影响
由表5可知,饲粮中添加不同碳链长度脂肪酸使乳脂中多种脂肪酸含量发生变化。SMCFA组 C12∶0、C14∶0、C14∶1、C16∶0 含量显著高于LCFA 组 (P <0.05),依 次 提 高 38.49%、38.53% 、47.83% 和 9.52%;同时亦显著高于对照组(P <0.05),依次提高 17.25%、26.85%、27.10% 和 7.50%。乳脂中 C18∶0、t-9-C18∶1、t-11-C18∶1、c-9-C18∶1、共轭亚油酸(CLA)含量LCFA组则显著高于SMCFA组(P<0.05),分别提高 29.49%、71.43%、36.28%、17.86% 和25.93%,但除t-9-C18∶1含量外,与对照组均差异不显著(P >0.05)。乳脂中 C4∶0、C6∶0、C8∶0、C10∶0、c-9,c-12-C18∶2、C18∶3 和 C20∶0 含量各组间均差异不显著(P>0.05)。
由表5可知,饱和脂肪酸(SFA)、<C16含量LCFA组显著低于SMCFA组(P<0.05),分别降低6.60%和19.49%,其中<C16含量LCFA组亦显著低于对照组(P<0.05),降低幅度为8.70%。单不饱和脂肪酸(MUFA)、>C16含量SMCFA组则显著低于 LCFA组(P<0.05),分别降低16.91%和21.39%,其中 >C16含量对照组亦显著低于LCFA组(P<0.05),降低幅度为6.69%。各组间多不饱和脂肪酸(PUFA)含量没有显著差异(P >0.05)。
硬脂酰辅酶A去饱和酶,即Δ-9去饱和酶,能在脂肪酸碳链9号位置引入不饱和双键,是乳腺组织内源合成MUFA的关键酶。Δ-9去饱和酶活性可通过乳脂中 C14∶1/C14∶0、C16∶1/C16∶0、c-9-C18∶1/C18∶0 的比值来反映[7]。由表 6 可知,各组间Δ-9去饱和酶活性无显著差异(P>0.05)。
3 讨论
3.1 饲粮添加不同碳链长度脂肪酸对泌乳奶牛生产性能的影响
本试验模拟荷斯坦奶牛典型乳脂结构中SMCFA和LCFA的比例,对反刍动物乳腺2种摄取来源脂肪酸对乳脂合成的影响进行了研究。与对照组相比,SMCFA组乳脂率提高3.71%,LCFA组乳脂率降低10.51%。奶牛饲粮中富含长链饱和脂肪酸对乳脂率有提高作用[8],而大量研究表明添加富含长链多不饱和脂肪酸的油脂不会提高奶牛乳脂率,且会出现乳脂率降低情况[9-10],可能的原因:一是泌乳前期奶牛能量处于负平衡状态,添加油脂可提高乳产量,而乳产量与乳脂率成负相关效应,因此导致乳脂率下降;二是在乳腺中某些特定反式脂肪酸,如t-10,c-12-CLA,抑制乳脂合成;三是饲粮添加过量油脂使瘤胃内纤维分解菌活性受到抑制,乙酸比例下降,减少了乳腺合成乳脂的前体物。本试验LCFA组脂肪酸含有大量不饱和脂肪酸,其中MUFA较高,PUFA较低,表明饲粮添加富含MUFA的油脂对奶牛乳脂率亦有负面影响,这与Rego等[11]报道的结果一致。

表5 饲粮添加不同碳链长度脂肪酸对泌乳奶牛乳脂肪酸组成的影响Table 5 Effects of supplemental fatty acidsw ith different carbon chain lengths on fatty acid composition in m ilk of lactating dairy cows %

表6 饲粮添加不同碳链长度脂肪酸对泌乳奶牛乳腺Δ-9去饱和酶活性的影响Table 6 Effects of supplemental fatty acidsw ith different carbon chain lengths on Δ-9 desaturase activity in mammary gland of lactating dairy cows
3.2 饲粮添加不同碳链长度脂肪酸对泌乳奶牛乳脂肪酸组成的影响
大量研究表明,外源途径添加LCFA引起乳脂中LCFA比例升高,SMCFA比例相应降低[1]。在乳脂降低综合症情况下,奶牛乳脂中SMCFA比例显著降低,而LCFA比例没有受到影响[2]。本试验结果亦显示,饲粮添加LCFA显著降低SMCFA中C12∶0和C14∶0含量,其原因可能为从血液到达乳腺的LCFA含量升高,乳腺中乙酰辅酶A羧化酶mRNA表达量受到抑制,因此乳腺从头合成SMCFA能力降低。乳脂中SMCFA占总脂肪酸的40%~55%,对于乳甘油三酯结构的稳定性有很重要的作用[12]。
奶牛饲粮含有较少量的SMCFA,乳脂中SMCFA几乎全部是瘤胃发酵产生的乙酸、β-羟丁酸在乳腺组织从头合成而来的。本试验通过饲粮途径外源添加SMCFA,SMCFA作为SFA,在瘤胃中稳定性高于不饱和脂肪酸,进入小肠后,作为碳链较短的脂肪酸,被小肠吸收,迅速进入肝脏氧化供能[13],因此供应至乳腺的比例较少。随着碳链长度增加,供应至乳腺比例逐渐增加[14]。有研究报道,添加的SMCFA至乳腺的转化效率分别为0(C6∶0)、0(C8∶0)、33%(C10∶0)、67%(C12∶0)、83%(14 ∶0)和 58%(16 ∶0)[4],饲 粮 添加富含C12∶0和 C14∶0 的椰子油导致乳脂中 C12∶0 和C14∶0含 量 升 高[15]。在 本 试 验 中,SMCFA 组C4∶0、C6∶0、C8∶0 和 C10∶0 含量与其他 2 组相比没有显著差异,说明以上脂肪酸在没有到达乳腺之前即被肝脏吸收。
衡量乳腺Δ-9去饱和酶活性指标有C14∶1/C14∶0、C16∶1/C16∶0、c-9-C18∶1/C18∶0 和 c-9,t-11-C18∶2/t-11-C18∶1,其中 C16∶0、C18∶0、t-11-C18∶1均可通过体脂分泌或消化道吸收分泌至乳脂中,只有C14∶0完全通过乳腺组织从头合成,因此 C14∶1/C14∶0 最能代表 Δ-9 去饱和酶活性[7]。本试验中,添加SMCFA饲粮显著提高乳脂中C14∶0和 C14∶1 含量,但 C14∶1/C14∶0 无显著变化,说明从饲粮外源添加至乳腺的一部分C14∶0通过Δ-9去饱和酶去饱和为C14∶1,但这种去饱和作用对于Δ-9去饱和酶的活性没有影响。
4 结论
饲粮添加400 g/d SMCFA或LCFA使泌乳奶牛乳脂率和乳脂肪酸组成发生改变,添加LCFA有降低乳脂率的趋势,并降低了乳脂中SMCFA的含量;而添加 SMCFA提高了乳脂中 C12∶0和C14∶0含量,并且对乳脂合成的贡献高于LCFA。
[1] GLASSER F,FERLAY A,DOREAU M,et al.Long-chain fatty acid metabolism in dairy cow s:a meta-analysis ofm ilk fatty acid yield in relation to duodenal flow s and de novo synthesis[J].Journal of Dairy Science,2008,91(7):2771-2785.
[2] LOOR J J,HERBEIN J H.Exogenous conjugated linoleic acid isomers reducem ilk fat concentration and yield by inhibiting de novo fatty acid synthesis[J].The Journal of Nutrition,1998,128(12):2411 -2419.
[3] KALAC P,SAMKOVA E.The effects of feeding various forages on fatty acid composition of bovinem ilk fat:a review[J].Czech Journal of Animal Science,2010,55(12):521-537.
[4] KADEGOWDA A K,PIPEROVA L S,DELMONTE P,et al.Abomasal infusion of butterfat increases m ilk fat in lactating dairy cows[J].Journal of Dairy Science,2008,91(6):2370-2379.
[5] VYASD,TETER B B,ERDMAN R A.M ilk fat responses to dietary short and medium chain fatty acids in lactating dairy cows[J].Journal of Dairy Science,2010,93(E-Suppl.1):444.
[6] BU D P,WANG JQ,DHIMAN T R,et al.Effectiveness of oils rich in linoleic and linolenic acids to enhance conjugated linoleic acid in m ilk from dairy cows[J].Journal of Dairy Science,2007,90(2):998-1007.
[7] PETERSON D G,KELSEY J A,BAUMAN D E.Analysis of variation in cis-9,trans-11 conjugated linoleic acid(CLA)inm ilk fatof dairy cows[J].Journal of Dairy Science,2002,85(9):2164-2172.
[8] WANG JP,BU D P,WANG JQ,et al.Effect of saturated fatty acid supplementation on production and metabolism indices in heat-stressedm id-lactation dairy cows[J].Journal of Dairy Science,2010,93(9):4121-4127.
[9] CHELIKANI P K,BELL J A,KENNELLY J J.Effects of feeding or abomasal infusion of canola oil in Holstein cows.1.Nutrient digestion andm ilk composition[J].Journal of Dairy Research,2004,71(3):279-287.
[10] 尹福泉,嘎尔迪,刘瑞芳,等.日粮中添加油料籽实对奶牛生产性能及乳脂脂肪酸组成的影响[J].动物营养学报,2008,20(3):261 -267.
[11] REGO O A,ALVES S P,ANTUNES L M,et al.Rumen biohydrogenation-derived fatty acids in m ilk fat from grazing dairy cow s supplemented w ith rapeseed,sunflower,or linseed oils[J].Journal of Dairy Science,2009,92(9):4530-4540.
[12] MOORE JH,CHRISTIEW W.Lipid metabolism in themammary gland of rum inantanimals[J].Progress Lipid Research,1979,17(4):347-395.
[13] LEVEILLE G A,PARDINI R S,TILLOTSON J A.Influence ofmedium-chain triglycerides on lipid metabolism in the rat[J].Lipids,1967,2(4):287 -294.
[14] GRUMMER R R,SOCHA M T.M ilk fatty acid composition and plasma energy metabolite concentrations in lactating cows fedmedium-chain triglycerides[J].Journal of Dairy Science,1989,72(8):1996 -2001.
[15] HRISTOV A N,VANDER POL M,AGLE M,et al.Effect of lauric acid and coconut oil on rum inal fermentation,digestion,ammonia losses from manure,and m ilk fatty acid composition in lactating cows[J].Journal of Dairy Science,2009,92(11):5561-5582.
