牛瘦素受体基因表达及其结合瘦素特性研究
2011-03-10李林强田万强昝林森
李林强,田万强,昝林森
(1.陕西师范大学食品工程与营养科学学院,陕西 西安 710062;2.杨凌职业技术学院动物工程系,陕西 杨凌 712100;3.西北农林科技大学动物科技学院,陕西 杨凌 712100)
牛瘦素受体基因表达及其结合瘦素特性研究
李林强1,田万强2,3,昝林森*3
(1.陕西师范大学食品工程与营养科学学院,陕西 西安 710062;2.杨凌职业技术学院动物工程系,陕西 杨凌 712100;3.西北农林科技大学动物科技学院,陕西 杨凌 712100)
[目的]以秦川牛为研究对象,克隆、原核表达秦川牛瘦素受体(Lepr)基因的蛋白功能区,分析表达蛋白对瘦素结合特性,为研究瘦素受体与瘦素结合的调节机制提供理论基础。[方法]本研究通过RT-PCR法从秦川牛肉m RNA扩增获得Lepr IG基因的cDNA序列,克隆至p MD18-T载体获得重组质粒,并进行序列测定。将测序正确的cDNA序列定向克隆到p ET 30a(NdeI/XhoI),构建表达载体,并转化BL21(DE3)大肠杆菌,IPTG诱导后进行SDS-PAGE分析,镍柱亲和层析法分离纯化融合蛋白,高效液相色谱—迎头法分析重组表达蛋白对廋素结合功能。[结果]表明秦川牛Lepr IG基因在大肠杆菌中获得高效表达;Lepr IG融合蛋白以可溶性和包涵体2种形式表达;高效液相色谱—迎头分析法进一步表明重组表达蛋白具有结合瘦素的功能。[结论]秦川牛Lepr IG基因在大肠杆菌中获得高效表达,表达蛋白具有结合瘦素的功能,为通过调节瘦素受体对瘦素结合,抑制能量代谢,促进秦川牛脂肪沉积奠定了基础。
牛;瘦素受体;IG 基因;克隆;表达
Leptin具有参与调节摄食、能量代谢等功能,通过其受体(Leptin receptor,Lepr)发挥调节作用。因此,瘦素受体在调节体脂平衡和能量代谢等方面发挥着重要作用。Tartaglia等(1995)首先发现存在于小鼠脉络丛内的leptin受体(OB-R),并测得小鼠的OB-R m RNA呈单个带状,长度约 5.1 kb[1]。OB-R是一种跨膜受体,由细胞外的配体结合区、跨膜区及胞内区三部分组成[2]。小鼠的OB-R由894个氨基酸残基组成,人的OB-R由1165个氨基酸残基组成。在小鼠中 Leptin受体存在OB-Ra、OBRb、OB-Rc、OB-Rd和OB-Re 5种异形体(isoform),其中只有OB-Rb属于长受体,具有信号转导功能,为功能受体,其它的均为短受体。长受体与短受体的胞外长度一致,它们之间的差异在于胞内结构域的长度不同。这些异形体的胞外结构域、跨膜结构域及胞内结构域中头29个氨基酸序列完全相同[3]。OB-Rb的胞内区含有2个结构域,一个可以激活Janus激酶(Just another kinase或Janus kinase,JAK),另一个可以和转录蛋白的激活因子(signal transducer and activator of transcription,STAT)相互作用,借以调节转录,OB-Rb是唯一能进行信号转导并调节热量摄入和能量消耗的异形体[4]。目前研究最多的是OBRa,其胞内结构域由33个氨基酸组成,它不能激活JAK激酶,也不能激活STA Ts。因此,它不具有信号传递功能,它可能作为Leptin的结合蛋白,调节游离的 Leptin浓度或作为 Leptin转运蛋白,使Leptin经过血脑屏障到达其作用位点[4,5]。OB-Re是唯一的可溶性受体,其功能是作为转运蛋白,OB2Rc和OB2Rd的功能目前尚不清楚。
目前,对瘦素、瘦素受体基因及其功能的研究主要集中于人和小鼠,主要是为了研究人类肥胖疾病产生的机理、预防和治疗方法,对黄牛OB-R研究尚处初级阶段,本文以中国五大黄牛之一的秦川牛为研究对象,克隆表达OB-R基因的蛋白结合区,为调控OB-R与Leptin的结合,抑制牛能量代谢,促进牛肌间脂肪沉积提供理论基础。
1 材料与方法
1.1 PCR扩增目的片段
采用T rizol法提取秦川牛肌肉组织基因组m RN A,DNA酶处理后,1%琼脂糖凝胶电泳检测核糖体RNA 28S和18S带的完整性(紫外分光光度计测定波长260 nm和280 nm处的OD值1.8~2.0)。用Revert Aid TM First Strand cDNA Synthesis Kit(Fermentas Life Science)反转录得到cDNA。RT-PCR获得目标基因,引物序列见表1。RT-PCR条件:95℃3 min,1个循环;94℃20 s;55℃20 s;72℃1 min 40 s,35个循环;72℃10min,1个循环。

表1 RT-PCR引物
1.2 克隆载体的构建
回收纯化 PCR产物,将获得的 1728 bp的Lepr基因cDNA序列连接 T-Vector(pMD18-T载体),4℃连接过夜,热击转化大肠杆菌DH5α,挑12个克隆进行PCR,然后进行质粒抽提,酶切鉴定插入片段。

表 2 p ET 30a酶切体系

表3 Lepr基因和p ET 30a(Nde1/Xho1)连接体系
1.3 表达载体的构建以及在大肠杆菌BL21(DE3)中的表达
将阳性克隆及表达载体pET 30a质粒经NdeI和XhoI双酶切(酶切体系见表2)后,回收1728 bp的片段以及载体大片段,将目的片段和载体大片段连接(连接体系见表3)过夜后转化BL21(DE3)感受态细胞。菌液PCR初步鉴定正确后,提取质粒,用NdeI和XhoI鉴定正确后测序。挑取测序正确的单克隆于5 mL含100m g/L氨苄的 LB液体培养基中37 ℃恒温振荡培养 12~14 h。次日,按1∶100扩大培养至OD值为0.6时,加IPTG至终浓度为1 mmol/L,继续诱导培养3 h,同时将空载体p ET 30a的诱导表达作阴性对照。在不同的时间点收集菌液,以10000 r/min速度离心10min沉淀菌体,加入SDS蛋白上样缓冲液混匀后,煮沸10min,离心取上清10μL进行SDS-PAGE。
1.4 目标蛋白亲和纯化(His-tag亲和柱)
利用His-tag亲和柱进行目的蛋白的纯化。
1.5 Lepr蛋白与Leptin结合作用高效液相色谱—迎头分析法(high performance liquid chromatography-frontal analysis,HPLC-FA)
高效液相色谱一迎头分析法(HPLC-FA)是基于凝胶过滤迎头分析法的原理发展起来的色谱方法,适宜高亲和势的蛋白结合作用的分析,样品可不经任何处理直接进样分析,通过柱切换和手性色谱柱相连,可进行蛋白结合作用的立体选择性研究。
色谱条件:日本分光公司JascoUV-975紫外可见分光光度检测器;色谱柱为Pinkerton GFF II-S5-80内表面反相(IS RP)柱,150mm×4.6 mm ID,5 μm(Regis Chemical Company,Illinois,USA);流动相为67 mmol·L-1磷酸盐等渗缓冲液(p H 7.4,I=0.17 mol·L-1);流速0.2 m L·min-1;紫外检测波长290 nm;进样量900μL;柱温37℃。
样品溶液的配制:吸取500μL Leptin(0.1 ng/μg)溶液于容量瓶中,然后加入等体积的Lepr蛋白溶液(0.12 mg/mL),加入5 mL等渗磷酸盐缓冲液,摇匀,37℃温育1 h。以Leptin和Lepr蛋白溶液为对照。
2 结果与分析
2.1 RT-PCR克隆秦川牛Lepr产物电泳检测
RTμPCR克隆秦川牛Lepr产物电泳检测结果见图1。由图1可见所获目的基因在1750 bp附近,与预测结果一致。由图2可见测序结果与NCBI原序列比较同源性达99.77%。

图1 RT-PCR克隆秦川牛Lepr产物电泳检测
2.2 酶切鉴定插入片段鉴定
连接产物转化大肠杆菌后,选择白色菌落进行PCR,提取阳性菌落质粒用NdeI和XhoI进行双酶切鉴定,结果见图 2。结果表明 Lepr基因插入pMD18-T克隆载体。

图2 牛Lepr原核表达质粒双酶切鉴定
2.3 表达检测及高表达菌株筛选
将转化 pMD18-T和p ET 30a的BL-21(DE3)阳性克隆菌液在37℃、1 mM IPTG诱导3 h后收集菌体,进行SDS-PAGE电泳,结果见图3。由图3可见克隆得到了约66 k D的预期蛋白特异带,与理论估计的相对分子质量相符。表明菌株诱导后都有明显的目标蛋白Lepr表达(箭头标记处)。
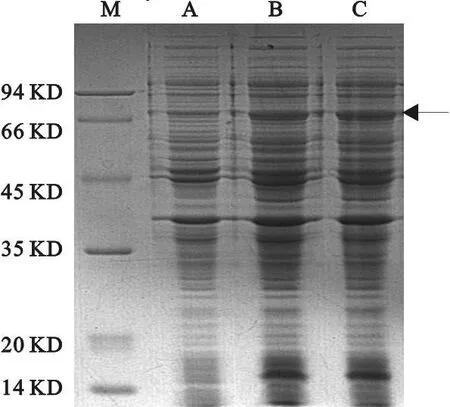
图3 表达检测及高表达菌株筛选
2.4 目标蛋白亲和纯化(His-tag亲和柱)
将菌体悬浮于PBS溶液中反复冻融3次,每克菌体加入80μL 10mg/L的溶菌酶及 40μL 100 nml/LPMSF(蛋白酶抑制剂)混匀,4℃放置30min后超声波破碎菌体,然后4℃8000×g离心15 min,分别收集上清和沉淀(沉淀主要为包涵体)进行SDS-PAGE。结果表明融合蛋白在上清中表达,也以包涵体的形式表达,不过上清中的表达量要大于包涵体,即融合蛋白可溶性表达大于包涵体表达(图4)。结果表明通过His亲和柱的分步纯化,可以从破菌上清中获得纯度合格目标蛋白Lepr(泳道F,箭头标记处)。

图4 目标蛋白亲和纯化
2.5 Lepr蛋白与Leptin结合作用HPLC-FA分析
Lepr蛋白与Leptin 37℃温育1 h经过HPLCFA,结果表明产生了新物质。结果提示Lepr蛋白与Leptin发生了一定程度的结合,进一步表明克隆表达的Lepr的蛋白结合区具有结合Leptin的功能。
3 讨论
Kielar等(1998)报道在下丘脑腹内侧核、侧部及弓状核存在有丰富的Leptin受体,在其它部位,如心 、胎盘 、肝 、肾 、胰 、脾 、肌肉 、胸腺 、前列腺 、睾丸 、卵巢、小肠、结肠、肾上腺中均有表达,但水平较低[6],与本文 Lepr组织表达特性研究结果基本相同。Houseknecht等(1996)报道,分泌到血液中的Leptin大部分通过与血清蛋白结合来运输,而后与下丘脑的Leptin受体结合,通过信号转导途径而发挥其生物学效应[7,8]。Wabitsch等(1996)也认为leptin与位于下丘脑及脂肪组织的Lepr结合,会产生抑制食欲、减少能量摄取、增加能量消耗、抑制脂肪合成的作用[9]。表明Lepr功能区表达重组蛋白与血清中leptin可能结合,进而可能抑制leptin与位于下丘脑Lepr的结合。
大理石花纹是评价牛肉品质的两个主要指标(大理石花纹和嫩度)之一。肌内脂肪组织比较丰富的牛肉在视觉上呈现大理石花纹状[10,11],而大理石花纹是日本[12,13],美国[14],韩国[15,16],中国等国牛肉品质分级的主要指标。因此,提高牛肉肌内脂肪含量,增加大理石花纹丰富度是肉牛生产研究的热点。本文克隆和表达Lepr蛋白结合区,结果表明该段基因可进行表达。通过HPLC-FA分析表达蛋白对Leptin的结合特性,结果表明Lepr功能区表达重组蛋白与 Leptin发生了一定程度的结合。这就为Lepr功能区表达重组蛋白对下丘脑Lepr与Leptin结合产生竞争性抑制,从而抑制牛能量代谢,促进牛肌内脂肪的沉积,以及为抑制牛能量代谢微生态制剂的开发提供一定的理论基础。当然,摄食和能量代谢的调节是多种神经递质在下丘脑互作的结果,因此,要阐明Lepr蛋白结合区对牛肌内脂肪调节机理还有许多要解决的问题。
[1]Tartaglia L A,Demb ski M,Weng X,et a l.Identification and expression cloning of a leptinrecep tor,OB-R[J].Cell,1995,83(7):1263-1271.
[2]M atsu ok a N,Ogaw a Y,Hosoda K,et a l.Human leptin receptor gene in obese Japanese subjects:evidence against eitherobesity-cau sing mutations or association of sequence variants with obesity[J].Diabetologia,1997,40:1204-1210.
[3]Ghilardi N,Skoda R C.T he Leptin receptor Activates Janus Kin ase 2 and Signals for Proliferation in a Factor-Dependent Cell Line[J].Molecular Endocrinology,1997,11(4):393-399.
[4]Ihle J N.Signaltran sducers and activators of transcription[J].Cell,1996,84:311-334.
[5]Reichlin S.Is Leptin a Secretion of th e Brain?[J].The Journal of Clinical Endocrin ology&Metabolism,1999,84(7):2267-2269.
[6]Kielar D,Clark J S C,Ciech anow icz A,et al.Leptin receptor is of ormsex pressed in human adipose tissue[J].Metabolism,1998,47(7):844-847.
[7]Houseknecht K L,Mantzoros C S,Kuliaw at R,et al.Evidence for leptin binding to proteins in serum of rodents and humans:modulation with obesity[J].Diabetes,1996,(45):1638-1643.
[8]Sinh a M K,Oh annesian J P,Heiman M,et a l.Nocturnal rise of leptin in lean,obese,and n on-in sulin-dependent diabetes mellitus subjects[J].Jou rn al of Clinical,1996,97(5):1344-1347.
[9]Wabitsch M,Jensen P B,Blum W F,eta l.Insulin and cortisol promote leptin production in cu ltured hum an fat cells[J].Diabetes,1996,(45):1435-1438.
[10]Woodw ard B W,Fernández M I.Comparison of conventional and organic beef production systems II.Carcass characteristics[J].Livestock Produ ction Science,1999,(61):225-231.
[11]Indurain G,Beriain M J,G oni M V,et a l.Com position and estimation of intramuscular and subcutaneous fatty acid composition in Spanish young bulls[J].Meat Science,2006,(73):326-334.
[12]JMGA.New Beef Carcass Grading Standards[R].Tokyo:Japan Meat Grading Association,1998.
[13]Yoshikaw a F,T oraichi K,Wada K,et a l.On a grading system for beef marbling[J].Pattern Recognition Letters,2000,(21):1037-1050.
[14]USDA.Official United States Standards for Grades of Carcass Beef[R].Washington,D C:Agricultural Marketing Service,1997.
[15]APGS.Report of Business for Animal Products Grading.Animal Products Grading System[R].Seoul:National Livestock Cooperatives Federation,1995.
[16]Kim C J,Lee E S.Effects of quality grade on the chemical,physical and sensory characteristics of Hanwoo(Korean native cattle)beef[J].Meat Science,2003,(63):397-405.
Studies on Protein Expression of Functional Region of Leptin Receptor Gene and its Character of Combinding Leptin in Cattle
LI Lin-qiang1,TIAN Wan-qiang2,3,ZAN Lin-sen*3
(1.College of Food Engineering and Nutritional Science,Shaanxi Normal University,Xi'an,Shaanxi 710062,China;2.Department of Animal Engineering,Yang ling Vocational&Technical College,Yangling,Shaan xi 712100,China;3 College of Animal Science and Tech no logy,North west A&F Universit,Yang ling,S haan xi 712100,China)
【Objective】In this paper,the functional region of leptin receptor IG gene was cloned and ex-pressed,the expressed protein combining leptin properties were analyzed,which would provide theoretical basis for the regulation of Lepr and leptin combinding.【Method】T he cDNA of Lepr IG gene was amplified from muscle m RNA of Qinchuan cattle by RT-PCR.The PCR product was cloned into the T vector pMD18-T to construct plasmid for sequencing.Then the cDNA was subcloned into the prokaryotic expressing plasmid vector pET 30a(NdeI/Xho1)and transformed into host Escherichia coli strain BL2l(DE3)for expression.The expression of Lepr function protein was induced by IPTG,and was identified by SDSPAGE.The protein was purified by Ni NTA column.Combinding leptin function of the expressed recombinant protein via high performance liquid chromatography frontal analysis(HPLC-FA).【Result】The results showed that Lepr IG gene was highly expressed in Escherichia coli,and the expression product was observed with soluble protein and inclusion body,the expressed recombinant protein had the function of combinding leptin.【Conclusion】In vitro expressed exogenous,Lepr IG gene protein had combinding leptin function and the study provided the foundation for inhibitting Qinchuan cattle energy metabolism and fat deposition through regulation of Lepr on combinding leptin.
Cattle;leptin Receptor;IG gene;Cloning;Expression
S823.2
A
1001-9111(2011)04-0001-05
2011-03-15
2011-04-22
陕西省13115科技创新工程(No.2010ZDGC-01),教育部长江学者与创新团队发展支持计划(No.IRt 0940)。
李林强(1971-),男,陕西扶风人,博士,研究方向为畜产品加工、动物资源开发与利用。
*通讯作者:昝林森(1963-),男,陕西扶风人,教授,博士生导师,研究方向为肉牛、奶牛遗传改良与生长发育调控。
