小鼠Aire逆转录病毒表达载体的构建及表达
2011-03-08李娟,张君,张毓
李 娟,张 君,张 毓
(1.天津医科大学免疫学教研室,天津300070;2.北京大学医学部免疫学系,北京100191)
Aire(autoimmune regulator)是一种自身免疫调节因子,其基因发现于1997年,由于Aire基因突变导致自身免疫性多内分泌病变-念珠菌病-外胚层营养不良(autoimmune polyendocri-nopathy candidiasis and ectodermal dystrophy,APECED),近几年引起人们的广泛关注。Aire基因主要表达于胸腺、淋巴结和脾脏,研究表明Aire通过诱导外周组织抗原(PTAs)在胸腺髓质上皮细胞中的表达促进自身反应性胸腺细胞的克隆清除,从而建立中枢耐受[1]。我们发现RelB缺陷小鼠和Aire缺陷小鼠中均存在CD4单阳性(single positive,SP)胸腺细胞发育缺陷[2],并且有研究表明RelB缺陷小鼠中Aire的表达显著减少[3],这些结果均提示Aire对SP胸腺细胞的最终成熟可能发挥重要作用。MigR1质粒是一种逆转录病毒载体,该载体多克隆位点MCS后加入内部核糖体进入位点IRES和绿色荧光蛋白GFP序列;逆转录病毒感染宿主细胞的范围较为广泛,且外源目的基因易整合入宿主细胞基因组,表达长期稳定[4]。本研究拟在MigR1的多克隆位点处插入小鼠Aire基因的ORF序列,构建能够稳定表达Aire蛋白的重组质粒,收集逆转录病毒上清,感染胸腺基质细胞系,为进一步研究Aire的生物学功能作准备。
1 材料与方法
1.1 材料
1.1.1 细胞株 293T细胞,MTEC9(小鼠胸腺基质细胞系,由北京大学医学部免疫学系T细胞研究室建立)。
1.1.2 质粒与菌株 MigR1逆转录病毒载体质粒、pHIT123质粒(编码鼠白血病病毒包膜蛋白)、pCGP质粒(编码鼠白血病病毒gag/pol)由陈友海教授(U- niversity of Philadelphia,USA)赠予;DH-5α感受态细菌由本室保存。
1.1.3 主要试剂 TRIzol®试剂购自Gibco BRL公司;Taq DNA聚合酶购自天为时代生物技术有限责任公司;BglII、XhoI等限制性内切酶、Pfu DNA聚合酶、逆转录试剂盒、Wizard Plus质粒提取试剂盒、CIAP(Calf Intestinal Alkaline Phosphatase)、NBT/ BCIP底物均购自Promega公司;质粒大量提取纯化试剂盒购自北京威格拉斯生物技术有限公司;Jet-PEITM转染试剂盒购自Polyplus公司;凝胶回收试剂盒(QIAEX II Gel Extraction System)购自Qiagen公司;anti-FLAG mAb、AP(碱性磷酸酶)-anti-rabbit-IgG购自Promega公司;anti-Aire多克隆抗体购自Upstate公司。
1.2 实验方法
1.2.1 Aire ORF扩增 利用Gene Runner软件分析Aire的核苷酸序列,在ORF两侧设计两条引物:上游引物:5’-GGG ATG GAA TGC TAC GCC-3’;下游引物:5’-TCA GGA AGA GAA GGG TGG TG-3’。以新鲜分离的小鼠胸腺基质细胞的cDNA为模板,Taq DNA聚合酶-Pfu DNA聚合酶联合使用进行PCR扩增,扩增aire全长ORF(94℃5min,94℃30s,55℃45s,72℃90s,72℃7min,共35个循环)。1.0%琼脂糖凝胶电泳确定目的片段。
1.2.2 Aire基因的克隆 利用QIAEX II Gel Extraction Kit凝胶回收并纯化目的片段,取纯化后产物5.0μL,加入10×PCR buffer1.0μL,dNTP 0.5μL,Taq DNA聚合酶0.5μL,无菌水3.0μL,混匀后72℃20min。取加A尾后的产物3.0μL加入2× rapid ligation buffer 5.0μL,pGEM-Teasy1.0μL和T4DNA ligase1.0μL,4℃连接过夜。用连接产物转化DH-5α感受态细菌,经蓝白斑筛选,挑取8~10个白色单克隆细菌,LA液体培养基振摇培养过夜,PCR鉴定(用克隆Aire ORF全长的引物),选取表达Aire的阳性克隆细菌,测序(北京三博远志生物技术有限公司)。将测序正确的阳性克隆菌液提取质粒备用。
1.2.3 Aire逆转录病毒表达质粒的构建及鉴定
1.2.3.1 BglII酶切位点的加入:在上下游引物的5’端分别合成BglII酶切位点,并在下游引物中加入FLAG标签,便于构建表达后的检测。
以1.2.2中提取的质粒为模板,使用Pfu DNA聚合酶进行PCR扩增,94℃5min,94℃30s,60℃45s,72℃90s,72℃7min,共35个循环。PCR产物胶回收、纯化、测序如前面所述。
1.2.3.2 MigR1质粒的单酶切以及去磷酸化:用BglII单酶切MigR1质粒(37℃,3~4h),凝胶回收酶切后的MigR1片段。取酶切后MigR115.0μL,加入10×buffer 5.0μL,0.01U/μL CIAP 5.0μL,无菌水25.0μL,37℃反应30min,在反应体系中再加入5μL 0.01U/μL CIAP,37℃,30min。使用QIAEX II Gel Extraction Kit将去磷酸化的产物立即进行脱盐和浓缩。
1.2.3.3 T-easy-Aire重组质粒的单酶切:用BglII单酶切T-easy-Aire重组质粒(37℃,3~4h),电泳,胶回收。
1.2.3.4 MigR1-Aire重组质粒的构建及鉴定:将酶切后T-easy-Aire 7.0μL和去磷酸化后MigR11.0μL混合,加入10×ligation buffer 1.0μL和T4DNA ligase1.0μL,4℃连接过夜。用连接产物转化DH-5α感受态细菌,经蓝白斑筛选,挑取10个白色单克隆细菌,LA液体培养基振摇培养过夜,提质粒,用BglII和XhoI进行酶切鉴定(BglII鉴定是否连接成功,XhoI鉴定连接方向是否正确),保存正向连接的菌种和质粒。选择正向连接的质粒,使用JetPEITM转染试剂盒转染293T细胞,转染48h后荧光显微镜观察转染效率。裂解细胞,Western blot检测Aire蛋白的表达。
1.2.4 逆转录病毒上清的收集 在10cm培养皿中接种包装细胞293T,细胞汇合度约20%,24h后将培养基替换为37℃预温的无血清DMEM,每孔10mL,在孵箱中放置10~30min;配制如下反应混合液:逆转录病毒质粒DNA 10μg,pHIT123质粒10μg,pCGP质粒10μg,2.5mol/L CaCl250μL,加无菌水补至500μL;将此混合液缓慢旋转加入500μL 2×HBS中,吹泡混匀,室温孵育30min;轻轻滴加至6孔板中,轻摇培养板混匀,37℃静置;5h后将培养孔中的液体换为37℃预温的DMEM-10%NCS,5~6mL/孔。48h后,收集上清,用0.45μm的滤器过滤或者1500r/min离心5min,获得的上清中含有短暂产生的病毒,将病毒上清分装,-70℃储存。
1.2.5 逆转录病毒上清体外感染胸腺基质细胞系
提前12~24h在6孔板中接种胸腺基质细胞系(大约2×105/孔);配制DMEM-10%NCS与polybrene (终浓度4μg/mL)的混合液;将6孔板中的培养基移去,在各孔中分别加入上述混合液与病毒上清(总体积2mL,可加入不同比例体积的逆转录病毒上清,但其体积应小于或等于感染混合液终体积的1/2),2500r/min离心90min;37℃孵育24h后将孔中液体更换为DMEM-10%NCS;48h后,用0.05%EDTA消化细胞,重悬在BSS-2%NCS中,制成单细胞悬液,FACS无菌分选GFP+细胞。裂解细胞,用anti-Aire polyclonal antibody进行Western blot检测。
2 结果
2.1 Aire ORF片段的获取 小鼠Aire基因全长1936bp,Gene Runner分析其ORF为58-1716,编码产物约60kDa。通过PCR扩增,得到约1.65kb的扩增片段(图1)。

图1 Aire ORF的扩增Fig 1 Amplification of Aire ORF
利用Gene Runner软件分析Aire基因的限制性酶切位点,未能找到适合的限制性酶切位点以便构建逆转录病毒表达质粒,因此我们设计了含BglII酶切位点的引物,并在下游引物中加入FLAG标签,便于构建表达后的检测。以Aire ORF为模板进行PCR扩增,得到含BglII酶切位点以及FLAG标签的Aire片段(Aire-BglII)(图2)。

图2 含BglII酶切位点及FLAG标签的Aire的扩增Fig 2 Amplification of Aire with BglII restriction enzyme cutting site and FLAG tag
2.2 Aire逆转录病毒表达载体的构建 经BglII酶切鉴定连接成功。用Gene Runner软件分析Aire序列的限制性位点,发现在473位有XhoI位点,若为正向连接,XhoI酶切后的小片段应在1243bp左右,若为反向连接,XhoI酶切后的小片段应在416bp左右。结果显示挑取的9个阳性克隆均连接成功,但是只有5个为正向连接(图3)。
选取正向连接的重组质粒转染293T细胞,转染48h后荧光显微镜观察转染效率约45%~50%(图4)。
利用anti-FLAG mAb进行Western blot检测,结果显示在60kDa左右有条带(图5),证明构建的逆转录病毒表达质粒能够成功表达Aire蛋白。
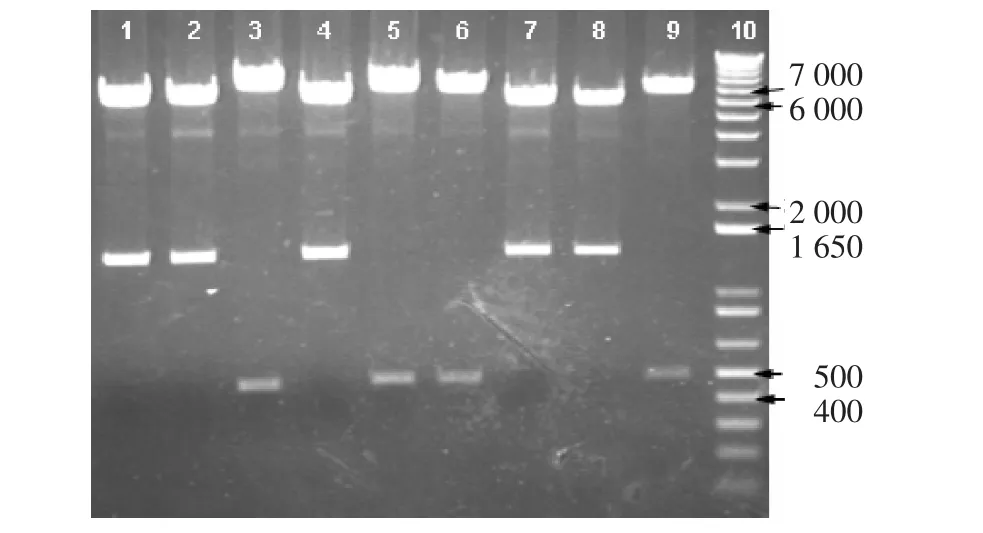
图3 MigR1-Aire酶切鉴定Fig 3 Verification of recombinant MigR1-Aire by restriction enzyme digestion

图4 转染后293T细胞的GFP表达Fig 4 The expression of GFP of 293T cells after transfection

图5 Western blot鉴定Aire蛋白的表达Fig 5 Western blot analysis of Aire protein
构建的MigR1-Aire示意图见图6。

图6 构建的MigR1-Aire示意图Fig 6 Sketch map of the recombinant plasmid MigR1-Aire
2.3 Aire蛋白在MTEC9细胞内强制性表达 用质粒MigR1-Aire及MigR1转染包装细胞系293T细胞,收集病毒上清感染MTEC9,流式细胞仪检测GFP的表达,感染效率约30%~35%(图7)。

图7 逆转录病毒上清感染MTEC9的效率检测Fig 7 The efficiency of infection with retroviral supernatant in MTEC9
MTEC9-Aire的裂解产物经抗体杂交后在60kDa左右出现条带,而MTEC9-MOCK则没有,表明Aire蛋白在MTEC9细胞内表达成功(图8)。

图8 Aire在MTEC9中的表达检测Fig 8 WesternblotanalysisofAireproteininMTEC9afterinfection
3 讨论
胸腺内T淋巴细胞的发育受到复杂而精细的调控,目前对于调控DN/DP/SP这几个阶段的转换及其中一些重要事件(如阳性选择、阴性选择等)的分子机制已有很多报道,但是关于SP胸腺细胞在髓质区分化过程的分子调控机制仍缺乏详细阐述。
我们对RelB缺陷小鼠和Aire缺陷小鼠的研究提示Aire在CD4SP胸腺细胞的最终成熟阶段可能发挥关键作用[2]。近几年的研究发现,Aire除了能够通过诱导PTAs的表达诱导中枢免疫耐受,还能通过参与调节胸腺髓质上皮细胞(medullary thymic epithelial cells,mTECs)的分化及功能的其它多方面来诱导中枢免疫耐受,如调节某些参与抗原提呈及处理的分子[5-6],调节mTECs的分化[7-8],调节mTECs中趋化因子CCR4和CCR7配体的表达[9]等。Aire究竟通过何种机制影响胸腺细胞的终末分化?要阐明此机制,构建能够融合表达GFP和Aire蛋白的表达质粒是非常有必要的。
我们可以分离Aire缺陷小鼠的mTECs,将其与正常小鼠来源的SP1细胞共育,检测其能否支持SP3向SP4的转化;若不能支持,我们将使用本研究中构建的Aire逆转录病毒表达载体,在Aire缺陷小鼠的mTECs中强制性表达Aire,观察能否纠正此缺陷。我们曾试图找到表达Aire的胸腺基质细胞系,但是在多株小鼠胸腺基质细胞系中并没有找到表达Aire的细胞系。本研究利用收集的逆转录病毒上清感染胸腺基质细胞系MTEC9,在该细胞系中成功表达Aire,为进一步研究Aire的强制性表达对胸腺基质细胞诱导SP胸腺细胞分化功能的影响奠定了基础。
[1] Anderson MS,Venanzi ES,Klein L,et al.Projection of an immunol ogical self shadow within the thymus by the Aire protein[J].Science, 2002,298(5597):1395
[2] Li J,Li Y,Yao JY,et al.Developmental pathway of CD4+CD8-medullary thymocytes during mouse ontogeny and its defect in Aire-/-mice[J].Proc Natl Acad Sci USA,2007,104(46):18175
[3] Zuklys S,Balciunaite G,Agarwal A,et al.Normal thymic architecture and negative selection are associated with Aire expression,the gene defective in the autoimmune-polyendocrinopathy-candidiasis-ectodermal dystrophy(APECED)[J].J Immunol,2000,165(4): 1976
[4] Pear WS,Miller JP,Xu L,et al.Efficient and rapid induction of a chronic myelogenous leukemia-like myeloproliferative disease in micereceivingP210bcr/abl-transducedbonemarrow[J].Blood,1998, 92(10):3780
[5] Anderson MS,Venanzi ES,Chen Z,et al.The cellular mechanism of Aire control of T cell tolerance[J].Immunity,2005,23(2):227
[6] Gray D,Abramson J,Benoist C,et al.Proliferative arrest and rapid turnover of thymic epithelial cells expressing Aire[J].J Exp Med, 2007,204(11):2521
[7] Gillard GO,Dooley J,Erickson M,et al.Aire-dependent alterations in medullary thymic epithelium indicate a role for Aire in thymic epithelial differentiation[J].J Immunol,2007,178(5):3007
[8] Dooley J,Erickson M,Andrew GF.Alterations of the medullary epithelial compartment in the Aire-deficient thymus:Implications for programs of thymic epithelial differentiation[J].J Immunol,2008, 181(8):5225
[9] Laan M,Kisand K,Kont V,et al.Autoimmune regulator deficiency results in decreased expression of CCR4and CCR7ligands and in delayed migration of CD4+thymocytes[J].J Immunol,2009,183(12):7682
