毛乌素沙地樟子松和油松人工林光合生理特性
2011-02-11丁晓纲李吉跃张方秋刘育贤
丁晓纲,何 茜,李吉跃,张方秋,刘育贤
(1.北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083;2.广东省林业科学研究院,广州 510520;3.华南农业大学,广州 430070;4.绵阳市林业科技推广中心,四川 绵阳 621000)
在大多数干旱地区,经常出现高温、强辐射和土壤干旱等现象[1],不同生活型植物在长期进化中已逐渐形成了适应干旱环境的生理机制和形态特征[2]。例如,1年生植物具有随降雨而萌发的随机型生存策略[3];短命植物利用很短的生长期完成生活史[4];而多年生植物则通过调节自身生长速率[5]和生理过程[6]增强对胁迫环境的抵抗力。以往的研究集中在干旱[7-8]、强辐射[9-10]、高温[11]、严寒[12]等条件下植物光合与水分生理特征的变化规律,其中通过植物气体交换特点来探讨植物的光合作用和水分利用效率及其与环境之间的关系,已引起广大学者的关注和研究,国内外已有不少报道[13-14],但有关水分胁迫对植物光合作用和气孔导度的影响多是以盆栽植物幼苗作为实验材料在人为控制条件下进行的,对于植物在自然干旱气候条件下的光合作用气体交换特征及其动态变化的研究虽然有学者进行了报道[15,17],但是关于樟子松和油松在沙地生态系统中的生理生态特性研究相对较少。在自然条件下研究毛乌素沙地腹地樟子松和油松人工林,生长季叶片气体交换和生态机理,同时探讨影响樟子松、油松光合速率的内外因素,以期为今后的研究工作提供理论基础。
1 试验地概况
毛乌素沙地位于鄂尔多斯高原的中部,属于荒漠草原地带,包括内蒙古自治区伊克昭盟的南部,陕西榆林地区的北部和宁夏回族自治区盐池的东北部。大部分地区属鄂尔多斯高平原向陕北黄土高原的过渡地区。
试验地位于伊克昭盟乌审旗图克苏木境内的毛乌素沙地整治开发研究中心针叶树人工林地,位于北纬38°57′-39°61′,东经109°21′-109°17′,海拔高度1200 ~1350 m,研究中心深居内陆,属温带大陆性半干旱气候,年平均温度6.4℃,年均降水量360 mm,其中80%降水集中于6月中旬至9月中旬;年蒸发量2300 mm;年均风速3.3m/s,大风扬沙日数40~50d,沙暴日数16d,大风多集中于4-5月间。土壤以典型草原发育而成的栗钙土型土壤和风沙土为主。流动沙地上植被覆盖率小于5%,土壤无发育;固定沙地地表有厚约1cm苔藓结皮层,土壤剖面微显发育,植被由油蒿(Artemisia ordosica)、柠条(Caragana intermedia)、臭柏(Sabina vulgaris)等建群物种组成。半固定沙地土壤有沙质草甸土、沼泽草甸土、碱化沼泽草甸土等。主要植被的建群种有假苇 拂子茅(Calamagrostis pseudophragmites)、寸草(Carex stenophylloides)、芨芨草(Achnatherum splendens)、碱茅(Puccinellia tenuiflora)、马蔺(Iris lacteal var.chinensis)等,植被覆盖率达到40%~80%。
2 材料与方法
2.1 试验材料
调查材料为毛乌素沙漠研究中心院内樟子松和油松人工林,两个树种以相同密度(625株/hm2)相邻种植于同一片造林地内,立地条件相同,林龄同为22a。林地为栗钙土型土壤,林下无植被覆盖,枯落物层3~5 cm。实验材料选取了长势良好、无病虫害的油松和樟子松标准木各3株。樟子松、油松人工林生长调查见表1。由表1得知樟子松年树高平均生长量为46.2 cm,比油松年平均生长量(32.3cm)高13.9cm,比油松年生长量多生长43%。樟子松平均胸径为17.6cm,比油松(12.6cm)高5cm,比油松胸径多生长39.6%。

表1 樟子松、油松人工林生长调查(林龄22a)
2.2 试验方法
2005年5月至9月中旬,选择晴朗无云的天气,连续3d测参试树种气体交换参数日变化,测定时选择待测样株树冠中上部外围向阳新梢上的倒3、4位的功能叶,测定时间为北京时间6:00-18:00,用枝剪剪下后迅速进行离体气体交换的重复观测,每个针叶枝每次连续采取9个稳定的数据,取其平均值。所有观测均在自然条件下进行。测定指标包括:净光合速率、蒸腾速率、气孔导度、胞间CO2浓度、叶温、环境因子(气温、光合有效辐射、空气相对湿度、大气CO2浓度)以及叶片与空气之间的水蒸汽压。气孔限制值则按Berry和Downton的方法计算(气孔限制值=1-胞间CO2浓度/大气CO2浓度)[18],水分利用效率=净光合速率/蒸腾速率。根据鄂尔多斯地区乌审旗气象站1985-2005年气象资料统计4-5月降雨量只占全年11%,而一年当中该地区7-9月的降雨量占全年降雨量的大部分[19]。
2.3 数据分析
用SAS 6.12进行方差分析,多重比较采用DUNCAN法。
3 结果与分析
3.1 生长季樟子松和油松气体交换参数日变化的季节动态
3.1.1 净光合速率 从图1中可以看出,在生长季内毛乌素地区油松和樟子松的光合速率日变化都表现出一定的日变化规律,即在白天时较高,在早晚时较低。但在不同的标准日内,变化规律差异较大,其差异表现在净光合速率的日变幅和峰值出现的时间两个方面。
在水分比较充足的8月、9月,两树种净光合速率的绝对值和日变幅明显高于干旱的5月、6月。在整个生长季,油松和樟子松的光合速率日变化都呈“单峰”曲线(见图1)。
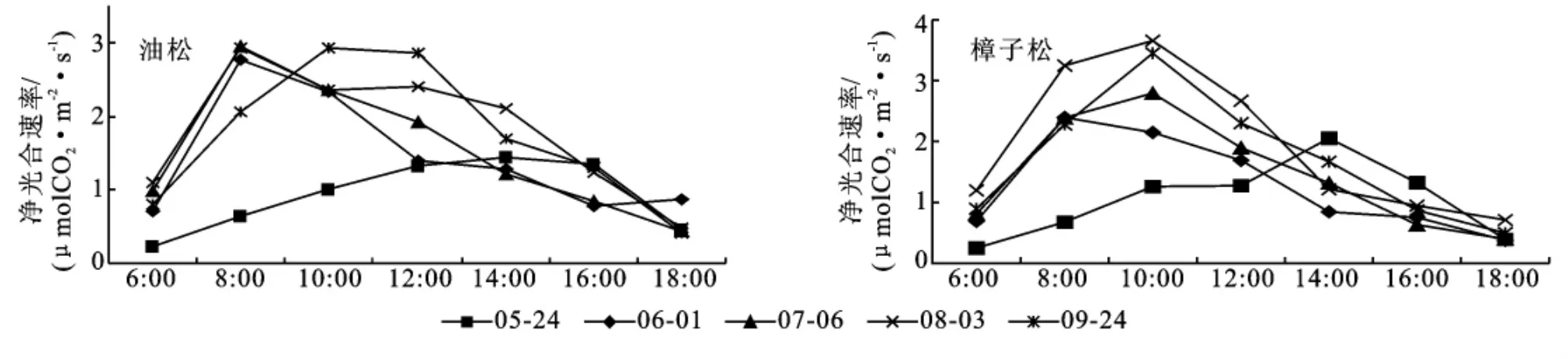
图1 生长季油松、樟子松人工林净光合速率日变化
3.1.2 蒸腾速率 叶蒸腾速率是从叶水平上表明植物的耗水强度。从图2可以看出,在生长季内,油松和樟子松的叶蒸腾速率基本上都表现出一定的日变化规律,在日出后,蒸腾速率开始增加,在正午前后达到最大,以后逐渐降低,到日落前后降至最低,但有时在16:00前后会稍有回升。在7月、8月、9月,林分土壤水分供给比较充足,标准日内叶蒸腾速率的峰值一般在出现在空气温度最高、湿度最低的中午前后,但在水分亏缺比较严重的4月、5月和6月初,标准日内叶蒸腾速率的峰值会提前出现.受水分供给状况和树木自身生长节律的影响,生长季内叶蒸腾速率的日变幅和绝对峰值表现出较大的季节差异,两树种蒸腾速率差异显著而且7-9月三个月明显大于4月、5月和6月初。
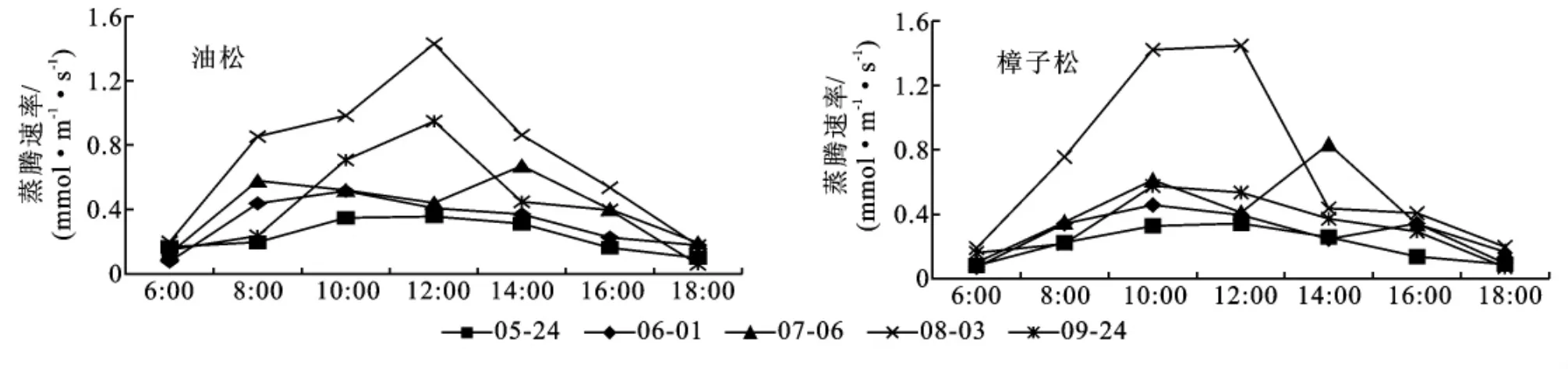
图2 生长季油松、樟子松人工林蒸腾速率日变化
3.1.3 水分利用效率 植物水分利用效率是指植物消耗单位水量所产生的同化物量,表示植物对水分的利用水平。这在环境差和水资源不丰富的条件下,对植物的初级生产力是非常关键的。单叶水平常用净光合速率和蒸腾速率之比来表示。从图3可以看出,上午时段的水分利用效率明显高于下午时段的水分利用效率,日变化呈“V”型曲线,其中5月24日两个针叶树种的最高峰值出现在下午16:00前后,9月24日两个针叶树种的最高峰值出现在上午8:00前后。该标准日内樟子松最高峰值为10.50μmolCO2/mmolH2O,油松最高峰值为8.65 μmollCO2/mmolH2O,在12:00前后,水分利用效率降至低谷。
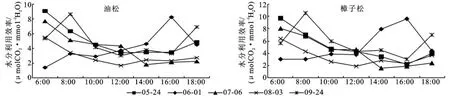
图3 生长季油松樟子松人工林水分利用效率日变化
根据各月份标准日内6:00-18:00的观测数据,处理后见表2。日平均净光合速率为该日测定净光合速率数据的平均值,日平均蒸腾速率与日平均水分利用效率由同样计算方法得出。方差分析表明两个针叶树种树日最高净光合速率值差异显著,尤其在8月樟子松日最高净光合速率为5.4μmol/(m2·s),比油松3.91μmol/(m2·s)高38.1%。两个针叶树种树日最高蒸腾速率值差异显著,樟子松显著低于油松,其中9月油松日最高蒸腾速率为1.34mmol/(m2·s)比樟子松0.64mmol/(m2·s)高109.4%。生长季除6月两树种水分利用效率相近以外,其它各月樟子松显著高于油松,其中9月樟子松水分利用效率为5.37μmolCO2/mmolH2O比油松4.03 μmolCO2/mmolH2O高33.2%。
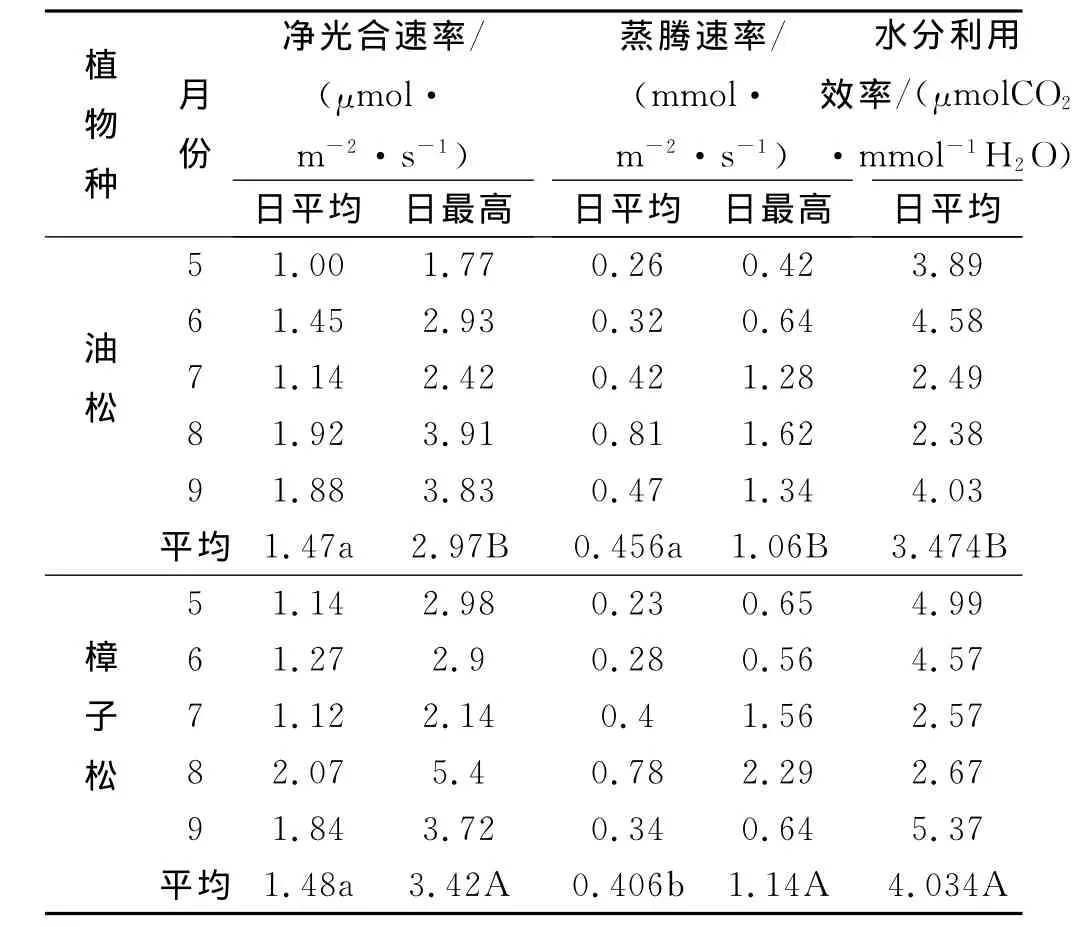
表2 油松和樟子松日变化光合速率、蒸腾速率及其水分利用效率平均值以及生长季平均值
3.2 大气因子对樟子松和油松气体交换的影响
光合作用是决定植物产量的最重要因素,不仅受到环境因子的影响,还受到植物本身诸多生理因子及其植物与环境相互作用的影响。各生理生态因子与净光合速率的相关系数见表3。从表3可知,生理生态因子与净光合速率之间的关系除了大气CO2浓度对Pn影响不显著以外,其他生理生态因子均显著影响着净光合速率。根据表3生理生态因子和Pn数据,取p=0.05,进行逐步回归分析方法,得到樟子松和油松两个回归方程(表4),从表4得知,限制油松、樟子松净光合速率的主要因子是光合有效辐射PAR和气孔导度。

表3 樟子松和油松的净光合速率与环境因子及其它生理变量的相关系数

表4 樟子松和油松净光合速率与生理生态因子之间的逐步回归方程
强烈的太阳辐射是引起一天中空气温度、相对湿度等一系列环境条件变化的根本原因。标准日中从8:00光合有效辐射开始持续上升,在10:00前后达到峰值然后逐渐下降,在18:00降到最低。气温的日变化从8:00起逐渐升高,于14:00前后达到峰值,之后依时序递降。随着光合有效辐射的增强及大气蒸发的加快,空气相对湿度急剧下降,净光合速率也迅速下降。大气因子对气孔导度的影响间接影响了植物光合作用。光对气孔导度的直接影响是短暂的,即在早晨随着光照的加强,刺激气孔开放,引起光合和蒸腾的迅速上升;而后太阳辐射的进一步增强使得气温上升及空气湿度下降,迫使气孔部分关闭,气孔导度下降明显;而后,气孔导度的变化趋势基本与湿度的变化一致,气孔的张开程度与湿度的关系更密切些,气孔导度与大气水汽压差呈相反的变化趋势,大气水气压差对应的高峰与气孔导度对应的低谷时间相一致,这与Buckley等研究气孔变化的结果相似,他们的观测结果显示水汽压差是气孔导度变化的主导因子[20]。
4 结论
(1)植物的光合作用、蒸腾作用、气孔导度和水分利用效率直接受太阳辐射和大气CO2浓度等环境因子的控制,同时它们也对环境变化具有很强的自我调节和适应能力,并做出相应的反应[21]。在自然条件下,植物总是通过气体交换与大气保持动态平衡,而光合作用是植物对环境变化很敏感的生理过程,植物适应环境是沿着有利于光合作用的方向发展。当环境条件如光照、温度超过其最适范围或生理忍耐限度时,植物通过调节气孔开度使Pn降低。毛乌素地区油松和樟子松的光合速率日变化都表现出一定的日变化规律,油松和樟子松的光合速率日变化都呈“单峰”曲线,但在不同的标准日内,变化规律的差异较大,表现在净光合速率的日变幅和峰值出现的时间两个方面。在水分比较充足的8月、9月,其净光合速率的绝对值和日变幅明显高于干旱的5月、6月份。
(2)油松和樟子松的叶蒸腾速率表现出一定的日变化规律,在日出后,蒸腾速率开始增加,在正午前后达到最大,以后逐渐降低,到日落前后降至最低,但有时在16:00前后会稍有回升。在7-9月三个月,林分水分供给比较充足,标准日内叶蒸腾速率的峰值一般出现在空气温度最高、湿度最低的中午前后,但在水分亏缺比较严重的4月、5月和6月初,标准日内叶蒸腾速率的峰值会提前出现。受水分供给状况和树木自身生长节律的影响,生长季内叶蒸腾速率的日变幅和绝对峰值表现出较大的季节差异,雨季明显大于旱季。
(3)6月、7月、8月针叶树水分利用效率曲线呈下降趋势,水分利用效率日变化特征为:上午时段的水分利用效率明显高于下午时段的水分利用效率。水分利用效率变化相对5月和9月稳定。针叶树水分利用效率日变化呈“V”型曲线,整个生长季日变化过程中,樟子松水分利用效率生长季日平均值为3.63 μmolCO2/mmolH2O,油松水分利用效率生长季日平均值为3.22μmolCO2/mmolH2O,樟子松的水分利用效率高于油松。生长季内樟子松各月的Pn、WUE均高于油松。表明油松属低光合、高蒸腾、低 WUE型的树种,樟子松属高光合、低蒸腾、高 WUE型的树种。
(4)在毛乌素沙地樟子松的净光合速率主要受到光合有效辐射、气孔导度和胞间CO2浓度的影响,限制油松净光合速率的主要因子是光合有效辐射和气孔导度。
[1]Etherington J R.Environment and Plant Ecology[M].Manchester:John Wiley & Sons,1975.
[2]Dodd M B,Lauenroth W K,Welker J M.Differential water resource use by herbaceous and woody plant life:forms in a short grass steppe community[J].Oecologia,2003,117:504-512.
[3]Volis S,Mendlinger S,Ward D.Adaptive traits of wild barley plants of Mediterranean and desertorigin[J].Oecologia,2002,133:131-138.
[4]Gutterman Y.Seed germination in desert plants[M]//Cloudsley-Thompson J L.Biotic Interactions in Arid Lands.NewYork:Springer-Verlag Berlin Heidelberg,1993.
[5]Heilmeier H,Wartinger A,Erhard M,et al.Soil drought increases leaf and whole-plant water use of Prunus dulcis grown in the Negev Desert[J].Oecologia,2002,130:329-336.
[6]Aranda I,Gill J,Pardos A.Effects of thinning in a Pinus sylvestris L.stand on foliar water relations of Fagus sylvatica L.seedlings planted with in the pine wood[J].Trees,2001,15:358-364.
[7]Batanouny K H.Plants in the Deserts of the Middle East[M].New York:Springer-Verlag Berlin Heidelberg,2001.
[8]Wicken G E.Ecophysiology of Economic Plants in Arid and Semi-arid Lands[M].New York:Springer-Verlag Berlin Heidelberg,1998.
[9]Peltier J P,Marigo D,Marigo G.Involvement of malate and mannitol in the diurnal regulation of the water status in members of Oleaceae[J].Trees,1997,12:27-34.
[10]Jiang G M,Zhu G J.Different patterns of gas exchange and photochemical efficiency in three desert shrub species under two natural temperature sand irradiances in Mu Us Sandy Area of China[J].Photosynthetica,2001,39(2):257-262.
[11]Snchez-Blanco M J,Morales M A,Torrecillas A,et al.Diurnal and seasonal osmotic potential changes in Lotus creticus palnts grown under saline stress[J].Plant Science,1998,136:1-10.
[12]Weiser C J.Cold resistance and injury in woody plants.Science,1970,169:1299-1278.
[13]李吉跃,Terence J B.多重复干旱循环对苗木气体交换和水分利用效率影响[J].北京林业大学学报,1999,21(3):1-8.
[14]李吉跃,张建国.北京主要造林树种耐旱机理及其分类模型的研究(Ⅰ):苗木叶水势与土壤含水量的关系及分类[J].北京林业大学学报,1993,15(3):1-11.
[15]Wartg H,Hart L,Zhou Z L,et a1.Dymumcal responses of Populus euphratica and Populus prunosa water potential to different depths of ground water level[J].Agricultural Research in the Arid Areas,2007,25(5):125-129.
[16]王海珍,韩路,周正立,等.胡杨、灰叶胡杨水势对不同地区水位的动态响应[J].干旱地区农业研究,2007,25(5):125-129.
[17]Li J,Zhang Q L,Gao R H,et al.Study on the population structure and distribution pattern of Populus euphratica populations in Ejina Oasi[J].Journal of Arid Land Resources and Environment,2008,22(2):187-190.
[18]Berry J A,Downton W J S.Environmental regulation of photosynthesis[M].New York:Academic Press,1982.
[19]丁晓纲,李吉跃,哈什格日乐.毛乌素沙地气候因子对樟子松、油松生长的影响[J].河北林果,2005,20(4):309-314.
[20]BuckleyT N,Mott K A.Dynamics of stomata water relations during the humidity response implications of two hypothetical mechanisms[J].Plane cell & Environment,2002,25(3):407-419.
[21]Yu G R,Zhuang J,Yu Z L.An attempt to establish a synthetic model of photosynthesis transpiration based on Stomata behavior formaize and soybean plants grown in field[J].Journal of Plant Physiology,2001,158:861-874.
