先进生物燃料导向的脂肪酸途径合成生物学改造
2011-02-09周雍进赵宗保
周雍进,赵宗保
1 中国科学院大连化学物理研究所,大连 116023
2 大连洁净能源国家实验室 (筹),大连 116023
3 中国科学院研究生院,北京 100049
先进生物燃料导向的脂肪酸途径合成生物学改造
周雍进1,3,赵宗保1,2
1 中国科学院大连化学物理研究所,大连 116023
2 大连洁净能源国家实验室 (筹),大连 116023
3 中国科学院研究生院,北京 100049
化石能源日益枯竭,迫切需要寻找新型燃料。脂肪族生物燃料由于其热值高、性能好而受到广泛重视。微生物脂肪酸代谢途径是生产先进生物燃料的重要途径。文中综述了近几年基于合成生物学理念改造脂肪酸途径的进展,介绍了合成生物学在微生物柴油、中长链脂肪醇、长链烃类化合物生物合成中的应用,并展望了脂肪族生物燃料的发展方向。
合成生物学,先进生物燃料,脂肪酸合成,生物柴油
燃料乙醇和生物柴油是当前生物燃料研究的热点,其生产成本在持续下降。但是燃料乙醇作为汽油替代品,其水溶性、挥发性、热值低等缺点制约着其大规模使用。先进生物燃料 (Advanced biofuel)是指以高产非粮作物、农林废弃物、其他可持续性生物质或者藻类为原料,通过化学、物理、生物甚至必要的组合技术,转化获得的具有与化石燃料相当甚至更高能量密度,易于储存和运输的燃料分子[3]。由于其性能优良和可持续性,先进生物燃料近来受到学术界和工业界重视。基于微生物代谢的生物技术对设计和制造先进生物燃料分子具有重要意义[4]。美国加州大学洛杉矶分校学者通过合成生物学手段改造大肠杆菌Escherichia coli氨基酸生物合成途径,合成了一系列碳链稍长的醇类化合物,并且大幅度提高了产量,为生物汽油发展提供了新思路[5]。生物柴油和石油来源的柴油性质相似,如果能进一步降低综合生产成本,将具有巨大市场潜力。然而,生物柴油当前面临最重要的瓶颈问题是植物油脂资源非常有限,难以支撑产业发展。另外,中长链 (碳链长度 C10-C18) 燃料如航空煤油目前尚没有非常有效的途径获取生物基替代品。最近关于微生物脂肪酸生物合成途径的认识和改造取得了一些成果,拓展了生物燃料的合成路线。
以下综述了近年来理性改造脂肪酸途径获取以脂肪酸代谢衍生物为目标的生物燃料研究,并展望了合成生物学策略在设计、改造生物燃料代谢途径中的应用。
1 合成生物学概念的内涵及其蓬勃发展
合成生物学一词早在1911年就出现在法国物理化学家 Stephane Leduc所著“生命的机理 (The Mechnism of Life)”一书中。1980年,Hobom B将其定义为利用重组 DNA技术对细菌进行基因工程改造[6]。随着分子系统生物学的发展,2000年Kool等在美国化学年会上重新提出,定义为基于系统生物学的遗传工程,从人工碱基DNA分子、基因片段、基因调控网络与信号传导路径到细胞的人工设计与合成[7]。西方学者对合成生物学比较一致的定义是:利用工程理念理性合成复杂、具有生物意义的不同层次系统—从单个生物分子到整个细胞、组织、器官,而这些合成的生物系统能执行自然界所没有的功能[8]。合成生物学、计算生物学与化学生物学一同构成系统生物技术的方法基础。
近几年合成生物学发展十分迅速,世界上多国政府部门及众多学者开始关注合成生物学研究,合成生物学也因此取得了蓬勃发展,并在天然产物、生物能源等领域取得了令人瞩目的成绩。比如:美国学者先后在 E. coli和酿酒酵母 Saccharomyces cerevisiae中构建了青蒿素前体生物合成途径[9-10],并进行代谢工程优化,在 S. cerevisiae中获得149 mg/L的青蒿酸[10],有望降低青蒿素生产成本,为非洲贫穷疟疾患者带来了希望。为了获得性能更好的汽油替代品,在 E. coli中引入 2-酮酸脱羧酶(KDC) 和乙醇脱氢酶 (ADH) 对氨基酸生物合成途径进行改造,构建了异源的醇类生物合成途径,合成了一系列碳链长度 (C3-C8) 醇类生物燃料,其中异丁醇产量最高达到22 g/L[5,11]。为了增加抗癌药物紫杉醇产量,降低其生产成本,在E. coli中构建紫杉醇前体生物合成途径,通过模块优化策略优化生物合成途径,其紫杉二烯产量达到1.0 g/L,5-羟基紫杉二烯产量达到 60 mg/L[12]。最引人注目的成果当属2010年Venter研究所成功地将人工合成的蕈状支原体 Mycoplasma mycoides基因组转入到山羊支原体Mycoplasma capricolum宿主细胞中,获得了具有生存和自我复制能力的新菌株[13]。
合成生物学的蓬勃发展使人们对微生物理性改造有了更深入的认识,也给现代工业生物技术提供了更有效的工具。近年来,生物燃料研究也充分利用了合成生物学最新发展理念,加速了其自身发展。除了上述利用合成生物学成功生产汽油替代品(C3-C8醇类) 外,人们开始利用合成生物学策略改造脂肪酸途径合成能量密度更高,性能更好的中长链生物燃料。
2 合成生物学改造脂肪酸途径合成生物燃料
模式微生物E. coli、S. cerevisiae等由于其遗传背景清楚,遗传操作手段成熟,被广泛用作合成生物学研究宿主。而脂肪生物合成能力较强的产油微生物遗传背景认识非常有限,尚难以进行合成生物学改造。目前脂肪族生物燃料的研究主要集中在E. coli中。
2.1 E. coli脂肪酸代谢途径
E. coli脂肪酸合成途径及其调控机制已得到广泛研究。如图1所示,乙酰-CoA经羧化得到丙二单酰-CoA后,经脂肪酸合成酶合成脂酰-ACP,随后经硫酯酶催化合成脂肪酸。在E. coli中,大部分脂酰-ACP经过甘油-3-磷酸酰基转移酶转移到甘油衍生物合成磷脂,少量的合成类脂等物质。当脂肪酸在E. coli中过量积累时,被快速转化为脂酰-CoA,然后经过β氧化途径分解成乙酰-CoA。E. coli脂肪酸生物合成途径受一系列机制严格调控,脂肪酸调节子的转录严格受到FadR和FabR调控,乙酰-CoA羧化酶和脂肪酸合成途径的FabH (3-酮酰-ACP合成酶III) 和FabI (烯酰ACP还原酶) 受到长链脂酰-ACP反馈抑制[14]。这些机制一起限制细胞内脂肪酸及其衍生物过量积累。所以,要大量合成脂肪类生物燃料,必须打破其调节机制,促进脂肪酸大量合成。最常用的策略之一是敲除脂肪酸分解途径关键基因fadD和fadE以减少脂肪酸分解;另外,过表达植物来源以及内源的硫酯酶解除长链脂酰-CoA反馈抑制,也可以加强脂肪酸合成[15]。
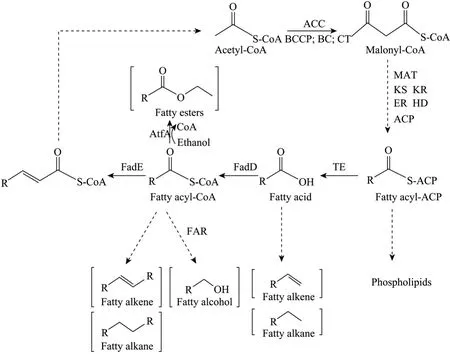
图1 E. coli中脂肪酸代谢途径[15]Fig. 1 The fatty acid metabolism in E. coli[15].
2.2 脂肪酸途径改造合成生物柴油
生物柴油是指短链一元烷醇脂肪酸酯,目前生物柴油通常是通过甘油三酯与短链醇类进行酯交换反应得到[16]。随着生物柴油需求增加,这个策略暴露了越来越多的缺点。首先其依赖于植物油脂供应,我国植物油脂资源极其短缺,现有油料作物年产油脂在1 000万t左右,尚不足以满足国内食用油消费市场需求[17]。近几年油脂进口量逐年递增,2009年超过900万t,是全球最大植物油净进口国。其次生物柴油转酯化需要消耗大量劳动力和能量,势必会减少其转化率,增加成本[2]。所以迫切需要寻找新技术以支撑生物柴油产业发展。为了解决这个问题,策略之一是寻找新的油脂来源。微生物油脂因其生产强度高,占用耕地面积少而受到广泛关注[18];另一种策略是利用微生物直接生产生物柴油,这种策略避免了转酯化等过程,也引起了重视。在 E. coli进行中过表达来源于运动单胞菌Zymomonas mobilis的丙酮酸脱羧酶增加乙醇供应,并表达来源于贝氏不动杆菌 Acinetobacter baylyi的酰基转移酶。在培养基中添加油酸进行有氧发酵,脂肪酸乙酯(FAEE) 产量达到1.28 g/L,FAEE含量达到细胞生物量的 26%[2]。由于这个过程要在胞外添加油酸,使其更像一个生物转化过程,但 FAEE高产量对未来脂肪酸途径改造研究具有重要参考价值。
为了实现生物柴油体内生物合成,敲除 E. coli DH1脂肪酸分解基因fadD增加脂肪酸供应,表达硫酯酶TesA、脂酰-CoA连接酶ACL、酯合成酶AtfA,从而构建了脂肪酸乙酯 (FAEE) 生物合成途径。然后过表达来源于 Z. mobilis的丙酮酸脱羧酶和乙醇脱氢酶加强乙醇生物合成途径,FAEE产量最大达到674 mg/L,为理论产量的7.4%[19]。相信对生物合成途径进行优化改造能进一步提高FAEE产量。
2.3 脂肪酸途径改造合成脂肪酸和脂肪醇
脂肪酸是微生物合成生物柴油的前体,也可以用作油脂化工行业的基本原料。如果能利用微生物过量生产脂肪酸,对生物燃料和生物基化学品生产具有重要价值。因此,已有研究对脂肪酸生物合成途径进行改造并增加脂肪酸产量。如通过敲除E. coli的fadD基因限制长链脂肪酸分解途径;过表达来源于油料植物种子的硫酯酶以加强脂肪酸合成速率;过表达乙酰-CoA羧化酶增加前体丙二酸单酰-CoA的供应;表达内源硫酯酶解除长链脂酰-CoA反馈抑制,最后脂肪酸产量达到2.5 g/(L·d),生产强度为0.024 g/(h·g DCW)[20]。在此基础上,将油料植物种子的硫酯酶和E. coli内源的硫酯酶在阿拉伯糖诱导启动子下串联表达,脂肪酸产量达到4.0 g/(L·d),生产强度达到0.04 g/(h·g DCW)。这已经很接近现在乙醇工业的生产强度0.1 g/(h·g DCW)[21]。有类似报道在敲除脂肪酸分解途径的菌株中过表达来源于植物的脂酰-ACP硫酯酶和乙酰-CoA羧化酶获得了约0.8 g/L脂肪酸[22]。在高产脂肪酸菌株的基础上,通过表达脂酰-CoA连接酶和脂酰-CoA还原酶,构建了脂肪醇生物合成途径,脂肪醇产量达到60 mg/L[19]。除E. coli外,也有报道对蓝细菌脂肪酸合成进行改造[23],在蓝细菌Synechocystis sp. PCC6803中过表达来源于好好芭 Simmondsia chinensis的脂酰-ACP还原酶,利用CO2作为碳源,脂肪醇产量达到0.2 mg/(L·OD730)。但是蓝细菌脂肪酸途经调节机理不够清楚,如何进一步进行代谢工程改造制约着产量进一步提高。
2.4 脂肪酸途径改造合成脂肪烃
烷烃和烯烃是汽油、柴油、航空煤油的最主要成分。在自然界中,许多生物能够体内能够合成烃类化合物,蓝细菌就是其中一种。然而,其生物合成的一些关键代谢步骤还没有得到清晰认识,因此也就很难通过合成生物学构建烃类生物合成途径。美国LS9公司通过差减策略从蓝细菌Synechococcus elongatus PCC7942中鉴定到烷烃生物合成基因脂酰-ACP还原酶编码基因和醛脱羰基酶基因,并对其表达酶生物化学性质进行了研究,其催化生物反应如图2所示。在E. coli中共表达脂酰-ACP还原酶和醛脱羰基酶,得到超过300 mg/L的烷烃,并且80%以上的烷烃是在细胞外。除了烷烃外,还生成了一定的醇类,这可能是由大肠杆菌内源的还原酶将中间产物脂肪醛还原产生的[24]。但是,在体外实验中脂酰-ACP还原酶和醛脱羰基酶共同催化脂酰-ACP并没有醇生成。如果增加醛脱羰基酶活力,并且通过代谢工程改造降低细胞内源的脂肪醛还原酶活力有可能将细胞内积累的醇类进一步转化为中长链烃,提高脂肪烃生产能力。
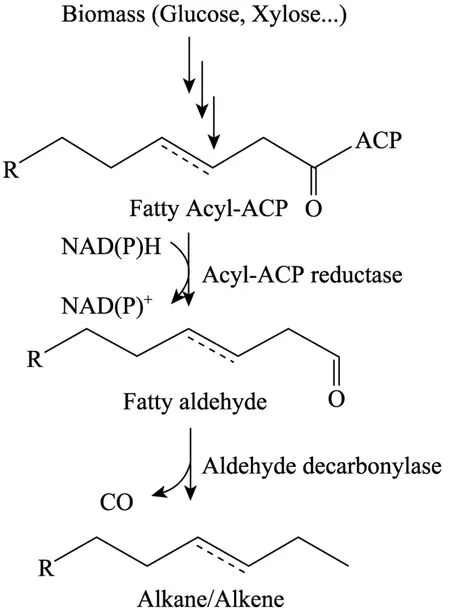
图2 脂肪烃生物合成途径Fig. 2 Alkane/Alkene biosynthesis pathway.
3 脂肪族生物燃料生物制造的展望
利用合成生物学理论与技术已实现了生物柴油、生物汽油、中长链烷烃等的微生物合成,但其产量还不够高,尚不能达到工业化生产水平。进一步通过合成生物学及代谢工程手段构建高产菌株是未来发展方向。将来可以从3个方面入手 (图3):1) 挖掘与改造合成生物学元件。自然界中资源丰富,有很多高效基因及酶有待于挖掘,设计高通量的筛选策略挖掘高效的基因元件为下一步合成生物学改造提供材料。近几年兴起的宏基因组技术和比较基因组学是筛选生物元件良好策略,如利用宏基因组技术从白蚁中发现了多个纤维素酶[25],最近又有报道利用宏基因组技术从牛胃中发现一系列纤维素酶[26],为纤维素高效利用菌株构建提供了材料。另外,自然界中挖掘的元件异源表达时有时候活性不高,选择性不够,需要进一步通过蛋白质工程和酶工程改造或者根据其功能设计新型元件使其更高效[27-28]。2) 组装生物燃料相关元件并协调其功能。DNA组装技术为代谢途径快速组装提供了很好的方案[29]。通过在E. coli中表达含有多种结构域的蛋白支架 (Protein scaffold),加州大学学者成功地将代谢途径相关的酶锚定在蛋白支架上,代谢流得到协调,目标产物生产能力提高77倍[30]。但是,蛋白质支架技术在其他宿主特别是真核生物以及其在不同途径中的有效性,目前尚不清楚,所以必须发展新技术调控代谢流。3) 提高宿主对燃料分子的耐受性。生物燃料分子的毒性直接影响细胞生长及代谢,从而限制产量进一步提高,所以增加宿主产物耐受性对获得高产很有必要。传统驯化筛选策略虽然能获得耐受性好的菌株,但比较耗时耗力,且菌株遗传性状不够稳定,易于产生回复突变。近几年发展的全局转录工程通过改造转录因子,获得了稳定的乙醇高耐受性菌株,值得借鉴[31]。另外,有学者通过人工转录因子技术,使E. coli对丁醇的耐受性提高到1.5%[32]。如果能够在这三方面整体突破,将会获得高产量脂肪类生物燃料生产菌株。
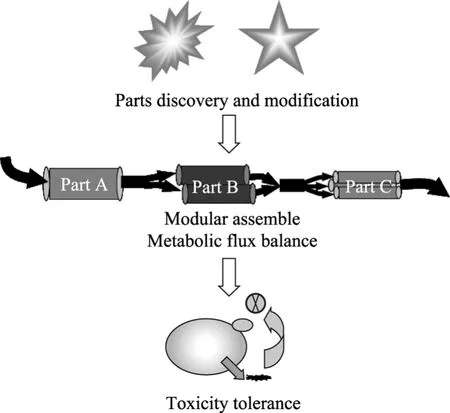
图3 脂肪族生物燃料合成生物学改造方向Fig. 3 The future aspects for enhancing fatty biofuel production.
目前,合成生物学宿主一般采用遗传操作平台成熟的模式生物。然而,它们的脂肪酸代谢能力还远低于产油微生物。另外,模式生物工程菌株可能存在如产物耐受性低、原料适应范围窄、抗逆性弱等性状缺陷。而产油微生物脂肪酸生物合成能力强,并且还存在高效异戊二烯途径,如果实现类胡萝卜素、甾醇等联产就有可能显著降低生物燃料成本[33]。因此,选择合适的宿主,构建其遗传操作平台,充分利用微生物代谢产物联产策略,可能是进一步提高微生物生物燃料生产竞争力的重要途径[34]。
总之,脂肪族生物燃料由于其热值高、不易挥发等良好性能受到广泛关注。但目前还面临菌株生产能力低、耐受性和原料适应性差等问题[34]。合成生物学理念和技术将促进脂肪族生物燃料生物制造快速发展,为生物能源产业提供技术支撑。
REFERENCES
[1] Kerr RA. Climate change. Global warming is changing the world. Science, 2007, 316(5822): 188−190.
[2] Kalscheuer R, Stölting T, Steinbüchel A. Microdiesel: Escherichia coli engineered for fuel production. Microbiology, 2006, 152(9): 2529−2536.
[3] What are advanced biofuel [EB/OL] ? [2011-04-07]. http://advancedbiofuelsusa. info/truly-sustainablerenewable- future.
[4] Peralta-Yahya PP, Keasling JD. Advanced biofuel production in microbes. Biotechnol J, 2010, 5(2): 147−162.
[5] Atsumi S, Hanai T, Liao JC. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature, 2008, 451(7174): 86−89.
[6] Hobom B. Surgery of genes. At the doorstep of synthetic biology. Medizin Klinik, 1980, 75(24): 14−21.
[7] Rawls RL. 'Synthetic Biology' makes its debut. Chem Eng News, 2000, 78(17): 49−53.
[8] Serrano L. Synthetic biology: promises and challenges. Mol Syst Biol, 2007, 3(1): 158.
[9] Martin VJJ, Pitera DJ, Withers ST, et al. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat Biotechnol, 2003, 21(7): 796−802.
[10] Ro DK, Paradise EM, Ouellet M, et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature, 2006, 440(7086): 940−943.
[11] Zhang KC, Sawaya MR, Eisenberg DS, et al. Expanding metabolism for biosynthesis of nonnatural alcohols. Proc Natl Acad Sci USA, 2008, 105(14): 20653−20658.
[12] Ajikumar PK, Xiao WH, Tyo KEJ, et al. Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science, 2010, 330(6000): 70−74.
[13] Gibson DG, Glass JI, Lartigue C, et al. Creation of a bacterial cell controlled by a chemically synthesized genome. Science, 2010, 329(5987): 52−56.
[14] Fujita Y, Matsuoka H, Hirooka K. Regulation of fatty acid metabolism in bacteria. Mol Microbiol, 2007, 66(4): 829−839.
[15] Liu TG, Khosla C. Genetic engineering of Escherichia coli for biofuel production. Annu Rev Genet, 2010, 44: 53−69.
[16] Tan TW, Lu JK, Nie KL, et al. Progress on biodiesel production with enzymatic catalysis in China. Chin J Biotech, 2010, 26(7): 903−906.谭天伟, 鲁吉珂, 聂开立, 等. 酶法合成生物柴油工业化研究进展. 生物工程学报, 2010, 26(7): 903−906.
[17] Zhao ZB, Hua YY, Liu B. How to secure triacylglycerol supply for Chinese biodiesel industry. China Biotechnol, 2005, 25(11): 1−6.赵宗保, 华艳艳, 刘波. 中国如何突破生物柴油产业的原料瓶颈. 中国生物工程杂志, 2005, 25(11): 1−6.
[18] Kosa M, Ragauskas AJ. Lipids from heterotrophic microbes: advances in metabolism research. Trends Biotechnol, 2011, 29(2): 53−61.
[19] Steen EJ, Kang YS, Bokinsky G, et al. Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature, 2010, 463(7280): 559−562.
[20] Lu XF, Vora H, Khosla C. Overproduction of free fatty acids in E. coli: implications for biodiesel production. Metab Eng, 2008, 10(6): 333−339.
[21] Liu TG, Vora H, Khosla C. Quantitative analysis and engineering of fatty acid biosynthesis in E. coli. Metab Eng, 2010, 12(4): 378−386.
[22] Lennen RM, Braden DJ, West RM, et al. A process for microbial hydrocarbon synthesis: overproduction of fatty acids in Escherichia coli and catalytic conversion to alkanes. Biotechnol Bioeng, 2010, 106(2): 193−202.
[23] Tan XM, Yao L, Gao QQ, et al. Photosynthesis driven conversion of carbon dioxide to fatty alcohols and hydrocarbons in cyanobacteria. Metab Eng, 2011, 13(2): 169−176.
[24] Schirmer A, Rude MA, Li XZ, et al. Microbial biosynthesis of alkanes. Science, 2010, 329(5991): 559−562.
[25] Warnecke F, Luginbuhl P, Ivanova N, et al. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature, 2007, 450(7169): 560−565.
[26] Hess M, Sczyrba A, Egan R, et al. Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science, 2011, 331(6016): 463−467.
[27] Leonard E, Ajikumar PK, Thayer K, et al. Combining metabolic and protein engineering of a terpenoid biosynthetic pathway for overproduction and selectivity control. Proc Natl Acad Sci USA, 2010, 107(31): 13654−13659.
[28] Zhang KC, Li H, Cho KM, et al. Expanding metabolism for total biosynthesis of the nonnatural amino acid L-homoalanine. Proc Natl Acad Sci USA, 2010, 107(14): 6234−6239.
[29] Shao ZY, Zhao H, Zhao HM. DNA assembler, an in vivo genetic method for rapid construction of biochemical pathways. Nucleic Acids Res, 2009, 37(2): e16.
[30] Dueber JE, Wu GC, Malmirchegini GR, et al. Synthetic protein scaffolds provide modular control over metabolic flux. Nat Biotechnol, 2009, 27(8): 753−759.
[31] Alper H, Moxley J, Nevoigt E, et al. Engineering yeast transcription machinery for improved ethanol tolerance and production. Science, 2006, 314(5805): 1565−1568.
[32] Lee JY, Yang KS, Jang SA, et al. Engineering butanol-tolerance in Escherichia coli with artificial transcription factor libraries. Biotechnol Bioeng, 2011, 108(4): 742−749.
[33] Zhao ZK, Hu CM. Progress of bioenergy-oriented microbial lipid technology. Chin J Biotech, 2011, 27(3): 427−435.赵宗保, 胡翠敏. 能源微生物油脂技术进展. 生物工程学报, 2011, 27(3): 427−435.
[34] Alper H, Stephanopoulos G. Engineering for biofuels: exploiting innate microbial capacity or importing biosynthetic potential? Nat Rev Microbiol, 2009, 7(10): 715−723.
Advanced biofuel-oriented engineering of fatty acid pathway:a review
Yongjin J. Zhou1,3, and Zongbao K. Zhao1,2
1 Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian 116023, China
2 Dalian National Laboratory for Clean Energy, Dalian 116023, China
3 Graduate University of Chinese Academy of Sciences, Beijing 100049, China
Biofuel is in high demand as an alternative energy source for petroleum and diesel. Fatty acid-based biofuel has higher energy density and better compatibility with existing infrastructures. Microbial fatty acid biosynthetic pathway is important to develop biofuel. In this article, recent progresses on the modification and reconstruction of fatty acid metabolism for the production of biofuel were reviewed, with a focus on micro-diesel, long chain fatty alcohol and alkane. Problems, solutions and directions for further development of fatty acid-based biofuel were also discussed in the respect of synthetic biology.
synthetic biology, advanced biofuel, fatty acid metabolism, biodiesel
随着经济快速发展,全球对能源需求剧增。传统化石能源日益枯竭难以支撑经济可持续发展,并且化石能源使用产生的温室气体对环境产生了巨大危害[1],所以迫切需要发展替代石化能源的可再生能源。与石化燃料相比,生物燃料源于可再生的生物质资源,其燃烧产生的CO2可经植物光合作用重新转化为生物质,实现绿色、可持续循环[2]。发展生物燃料对经济社会发展以及国家安全具有重要意义。得到了国内外学术界和政府部门的重视,取得了很多令人振奋的成果。
April 7, 2011; Accepted: June 10, 2011
Supported by:National Basic Research Program of China (973 Program) (No. 2011CB 707405).
Zongbao K. Zhao. Tel: +86-411-84379211; E-mail: zhaozb@dicp.ac.cn
国家重点基础研究发展计划 (973计划) (No. 2011CB707405) 资助。
