斜纹夜蛾核多角体病毒诱导甜菜夜蛾细胞凋亡的研究*
2011-01-24余倩
余 倩
(1.仲恺农业工程学院,广东 广州 510225;2.中山大学 有害生物控制与资源利用国家重点实验室,广东 广州 510275)
1972年Kerr等人[1]首次提出细胞凋亡的概念,即在一定的生理或病理条件下,细胞受内在遗传机制控制,自动结束生命的过程。细胞凋亡有明显形态学和生化学的特征,形态学特征包括:细胞核浓缩和片段化,胞质浓缩、细胞体急剧变小,细胞骨架解体,且常伴随有细胞质膜出泡现象[2]。生化学特征包括:染色质片段化,形成长度180~200 bp整数倍的寡聚核苷酸片段,琼脂糖凝胶电泳时呈现“梯状”条带 (DNA ladder)。
在多细胞生物体中,细胞凋亡作为一种抗病毒应答机制能限制入侵细胞中的病毒,从而减少子代病毒的产生,及造成流产性感染[3-5]。目前已发现多种杆状病毒可诱导昆虫离体细胞凋亡,如野生型苜蓿银纹夜蛾核多角体病毒诱导海灰翅夜蛾S.littoralis细胞 SL2 凋亡等[6]。
斜纹夜蛾 (Spodoptera litura,Splt)和甜菜夜蛾 (Spodoptera exigua,Se)同属夜蛾科灰翅夜蛾属的昆虫,亲缘关系很近,且斜纹夜蛾核多角体病毒 (Splt nucleopolyhedrovirus,SpltMNPV和SeMNPV基因组相似性很高[7-8],但这两种病毒不能交叉感染各自宿主。SeMNPV感染SpLi-221会引起细胞凋亡[9],而SpltMNPV感染Se301的研究还未见报导。本文在显微水平和亚显微水平上对SpltMNPV感染离体细胞Se301的过程进行了描述,并对感染失败原因进行了初步分析。
1 材料和方法
1.1 材料
1.1.1 昆虫细胞与病毒 野生型SpltMNPV中山大学分离株由本实验室保存。SeMNPV-SeUS1引自美国加州大学Riverside分校B.A.Federici教授实验室。甜菜夜蛾细胞Se301由荷兰Wageningen大学J.M.Vlak教授惠赠,用φ=10%胎牛血清的Grace's培养基(Invitrogen)27℃培养。
1.1.2 供试昆虫 斜纹夜蛾、甜菜夜蛾幼虫由本实验室养虫室提供,为人工饲料饲养的健康幼虫。人工饲料配方见参考资料[10],饲养温度27~28℃。
1.1.3 试剂 DAPI(4',6-Diamidine-2'-phenylindole dihydrochloride)购自Roche公司;细胞总DNA提取试剂盒购自Takara。
1.2 方法
1.2.1 病毒多角体扩增 将病毒多角体涂布于人工饲料表面喂饲四龄初昆虫幼虫,感染4~5 d后收集呈现典型病毒感染症状的幼虫 (虫体倒挂,身体肿胀,呈灰白色),置室温下静置2~3 d让其自然发酵。用10倍的PBS悬浮发酵的虫尸,经4层纱布过滤,滤液经3 000 r/min离心10 min,沉淀物重悬于适量PBS中,600 r/min离心5 min,收集上清。如此重复3~4次,直至多角体悬浮液呈乳白色,光学显微镜检查在多角体悬液中杂质很少,即为较纯的病毒多角体。
1.2.2 病虫血淋巴制备 用多角体喂饲四龄初幼虫,感染3 d后经镜检有极少量多角体出现即取血淋巴。用φ=70%乙醇将病虫表面消毒,剪开腹足,血淋巴滴入含w=0.1%苯基硫脲的Grace's培养基中,3 000 r/min离心10 min,除去血淋巴细胞,取上清经过滤灭菌保存,供细胞感染用。
1.2.3 病毒感染 在35 mm培养皿中以0.5×106个细胞/孔密度接种对数生长期的Se301细胞;待细胞贴壁1 h后,移出培养基,病毒以MOI为1感染细胞,27℃培养1 h并间隔摇动培养板;1 h后弃感染液,以无抗生素无血清Grace's培养基洗2次,然后加入Grace's培养基继续培养。加入病毒即开始计时,于感染后不同时间收集取样,3 000 r/min离心10 min,弃上清,细胞沉淀先放入液氮速冻30 min,再放入-70℃保存备用。
1.2.4 病毒滴度测定 采用终点稀释法对病毒进行定量测定[10],具体步骤参考 O'REILLY主编的《杆状病毒表达载体》。
1.2.5 电镜样品制备及观察 用病毒SeMNPV和SpltMNPV以MOI=1分别感染Se301细胞,于感染后0、24、48、72、96和120 h用细胞刮收集细胞,2 500 r/min离心5 min收集细胞。按常规电镜样品制备方法制备,简述包括前固定、漂洗、后固定、脱水、浸透、包埋、聚合、切片、染色步骤。镜检:用JEM-100CX型电镜在加速电压80 KV下观察。
1.2.6 DAPI染色 将固体粉末用双蒸水稀释成终质量浓度1~5 mg/mL作为储存液;用甲醇稀释储存液至终质量浓度1 μg/mL作为工作液。具体操作步骤参见Roche公司产品说明书。使用340 nm/380 nm滤片进行荧光显微镜观察。
1.2.7 细胞总DNA的提取 按照Merck公司的细胞总DNA提取试剂盒 (Suicide TrackTMDNA Ladder Isolation Kit)操作手册进行。
2 结果
2.1 SpltMNPV感染Se301细胞的光学显微镜观察SpltMNPV按照MOI=1的感染复数感染Se301
细胞,感染后每天用光学显微镜进行细胞观察。正常Se301细胞形态为长形梭状 (图1a);SeMNPV感染Se301细胞后细胞出现典型的感染症状,如细胞变圆;后期细胞中能观察到大量多角体 (图1b)。SpltMNPV感染Se301细胞光镜观察结果显示在感染后24~120 h期间细胞出现明显病理现象,且随着时间的延长病理症状越来越严重,在24~48 h时出现极少量凋亡小体,大部分细胞出现感染症状,如细胞变圆变大;也可观察到空泡细胞和少量细胞聚集 (图1c),到96~120 h时出现大量细胞聚集,空泡细胞明显增多,但始终观察不到病毒多角体 (图1d)。
2.2 SpltMNPV感染Se301细胞的电子显微镜观察

图1 病毒感染的Se301细胞光镜照片Fig.1 Light micrographs of Se301 cells infected

图2 SpltMNPV感染的Se301细胞电镜照片Fig.2 Electron micrographs analysis of Se301 cells infected with SpltMNPV
光镜观察显示感染细胞除产生少量凋亡小体外大部分细胞出现空泡和细胞聚集现象,为了更清楚准确的观察细胞内发生了什么病理变化,这些空泡细胞内到底有没有DNA存在,融合在一起的细胞是什么状态,我们收取SpltMNPV感染Se301细胞后24,48,72,96,120 h样品进行电镜观察,结果如图2显示:正常Se301细胞成单个长椭圆形,具有完整细胞器、细胞膜和核膜;细胞质和染色质分布均匀 (图2:a)。SeMNPV感染Se301细胞72 h整个细胞变成圆形,细胞质均匀分布,但细胞核膨大,病毒在核中央形成了病毒发生基质,再放大数倍后可观察到细胞核内有许多正在包装和已经包装好的病毒粒子 (图2:b,c)。SpltMNPV感染Se301细胞电镜观察显示:从感染后24~120 h均能观察到细胞病理现象,且晚期更剧烈。有些细胞只能看见细胞核膜包裹着高度凝聚的染色质,观察不到细胞膜和细胞质,在两个细胞间无清晰界限只有许多空泡存在;有一些细胞变圆细胞核膜完整且细胞核完好但细胞质中出现许多空泡;有一些细胞细胞质完全被泡状化且细胞核内染色质被高度浓缩凝聚并附于核膜周边 (图2:d,e,f);以上结果可被判断细胞出现了早期凋亡症状,但始终没有进行至细胞凋亡晚期出现凋亡小体。
2.3 DAPI染色
电镜结果表明细胞出现早期凋亡特征,为了更直观地观察细胞 DNA的变化,我们用 DAPI对SpltMNPV感染Se301细胞进行染色。DAPI是一种能与DNA强力结合的荧光染料﹐常被用于凋亡的检测;DAPI染剂能被紫外光激发,在荧光显微镜下观察为蓝色。
荧光显微镜观察结果如图3所示:正常Se301细胞由于细胞核内染色质分布均匀,DAPI染色后细胞核呈现为明亮而均匀的圆形 (图3:a);而被感染的Se301细胞从感染后24~120 h有明显变化(图3:b,c,d),例如有些细胞染色质分布不均,高度凝聚成不规则形状如月牙形,并集中在细胞核边缘;有些细胞细胞核染色荧光强度很弱,形状也不规则;还有些细胞中的DNA已经断裂,染色后呈现为许多个荧光小点并集中在核边缘上,细胞至感染后期也无太大改变并未见有大量凋亡小体产生。以上结果说明细胞出现了早期凋亡现象,而未进行至晚期,这些结果与电镜观察结果一致。
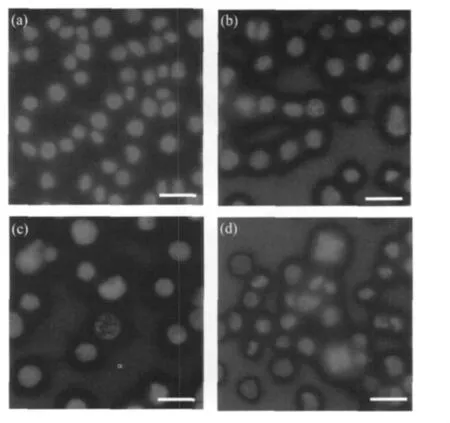
图3 DAPI染色的Se301细胞荧光显微镜观察Fig.3 Micrographes of Se301 Stained by DAPI
2.4 DNA ladder
细胞凋亡的另一个特征是出现DNA ladder现象,这属于凋亡晚期事件但比极晚期事件出现凋亡小体早。为了鉴定细胞凋亡进程是否进行至DNA ladder阶段,收取 SpltMNPV感染 Se301细胞24,48和72 h的细胞样品,提取其细胞总DNA进行电泳。实验结果如图4显示:SpltMNPV感染Se301细胞24,48和72 h样品与正常Se301细胞及SeMNPV感染Se301细胞72 h样品一样都没有出现典型梯度条带。这些结果说明SpltMNPV感染Se301细胞从24~72 h都未进行至DNA梯度条带的凋亡晚期阶段。

图4 Se301细胞总DNA的琼脂糖凝胶电泳分析Fig.4 Agarose gel electrophoresis of oligonucleosomal laddering isolated from Se301 cells
2.5 一步生长曲线测定
光镜检测一直未观察到SpltMNPV感染Se301细胞中有多角体产生,说明细胞的早期凋亡对病毒感染以及子代病毒产生具有严重影响。进行TCID50一步生长曲线测定检测病毒是否产生了BV。收取SpltMNPV感染Se301细胞24,48,72,96,120 h的细胞上清,同时也收取相应时间的SpltMNPV感染SpLi-221细胞作为阳性对照,用SpLi-221细胞测定SpltMNPV滴度。结果显示:SpltMNPV感染SpLi221细胞后病毒产生了大量有感染性的子代病毒,且感染后72 h达到最大值;而SpltMNPV感染Se301细胞样品中检测不到有感染性的BV产生。

图5 SpltMNPV感染SpLi-221或Se301细胞的BV增长曲线Fig.5 The BV replicative curves generated from an infection of SpltMNPV in Se301 or SpLi-221 cells
3 讨论
本研究通过光学显微镜和电子显微镜对SpltMNPV感染非受纳细胞Se301进行形态观察,以及DAPI染色等检测,首次发现被SpltMNPV感染的Se301细胞发生了早期凋亡但未能进行至晚期凋亡出现凋亡小体,本结果又一次证明了细胞凋亡是宿主抵抗病毒感染的防御机制,具有普遍的生物学意义。先前已有关于杆状病毒诱导离体细胞凋亡的报道:如野生型AcMNPV感染的SL2[6],美国白蛾核多角体病毒 (HyphantriacuneaNPV,HycuNPV)感染的Ld652Y细胞[11],但这些研究中细胞凋亡都进行至凋亡晚期即形成凋亡小体。杆状病毒诱导早期细胞凋亡是否影响病毒复制周期的完成至今还少有报道,本研究发现正是由于Se301发生了早期细胞凋亡从而阻止SpltMNPV产生BV和ODV子代病毒,可称Se301细胞为SpltMNPV的非受纳细胞。
对于任何一种病毒来说它进入宿主细胞和组织的能力和它在里面复制并释放有感染活性病毒的能力决定着这种病毒的宿主范围。SpltMNPV感染Se301细胞后在早期24 h时有极少数细胞产生凋亡小体,推测可能是病毒BV在病毒表面的糖蛋白GP64或其同源蛋白LD130参与下通过包内吞作用进入细胞后[12],其早期事件如早期基因的表达诱导了细胞凋亡。有研究已证实杆状病毒诱导的细胞凋亡可由感染早期事件所引起[13]。ie1是病毒感染的极早期基因,先前研究表明它能引起由AcMNPV诱导的Sf21细胞凋亡[14]。
被SpltMNPV感染的Se301细胞绝大部分只出现了早期凋亡并停滞在这个阶段,没有进行至晚期出现凋亡小体。推测病毒进入细胞后虽然病毒的早期事件引起了细胞凋亡的开启,可能病毒的抗凋亡基因Splt-iap4或Splt-p49得到了表达且具有抗凋亡功能,从而阻止了细胞凋亡的进程。早期基因表达后病毒借此机会进行了自身的复制和转录,DNA复制后病毒随即进行晚期或极晚期基因表达[15],由于某种未知原因,病毒晚期基因的表达可能受到抑制以至病毒不能完成BV的装配和产生子代病毒,所以在实验中我们观察到了以上的现象。
根据凋亡形态学的特征,本研究中的现象除一些与早期凋亡一致外,还有一些与凋亡特征不符,如细胞聚集融合在一起。由于受死亡信号刺激,典型的凋亡早期细胞应出现细胞膜卷曲皱缩,表面微绒毛消失,连接消失,与周围的细胞脱离等症状,而所观察到的现象却是几个细胞簇集在一起分不清细胞边界。实验中我们注意到,Se301细胞被SeMNPV感染后早期会出现细胞聚集现象,病毒产生子代病毒后又慢慢自行分开,所以推测以上现象为早期凋亡和病毒感染细胞双重影响的结果。
致谢本研究在中山大学庞义教授和杨凯教授的指导和帮助下完成,谨以感谢。
[1]KERR J F,WYLLIE A H,CURRIE A R.Apoptosis:a basic biological phenomenon with wide-ranging implications in tissue kinetics[J].Br J Cancer,1972,26:239-257.
[2]WYLLIE A H,KERR J F,CURRIE A R.Cell death:the significance of apoptosis[J].Int Rev Cytol,1980,68:251-306.
[3]ZHANG P,YANG K,DAI X,et al.Infection of wildtype Autographa californica multicapsid nucleopolyhedrovirus induces in vivo apoptosis of Spodoptera litura larvae[J].J Gen Virol,2002,83:3003 -3011.
[4]CLARKE T E,CLEM R J.In vivo induction of apoptosis correlating with reduced infectivity during baculovirus infection[J].J Virol,2003,77:2227 -2232.
[5]FENG G,YU Q,HU C,et al.Apoptosis is induced in the haemolymph and fat body of Spodoptera exigua larvae upon oral inoculation with Spodoptera litura nucleopolyhedrovirus[J].J Gen Viro,2007,88:2185 -2193.
[6]GERSHBURG E,RIVKIN H,CHEJANOVSKY N.Expression of the Autographa californica nuclear polyhedrosis virus apoptotic suppressor gene p35 in nonpermissive Spodoptera littoralis cells[J].J Virol,1997,71:7593-7599.
[7]PANG Y,YU J,WANG L,et al.Sequence analysis of the Spodoptera litura multicapsid nucleopolyhedrovirus genome[J].Virology,2001,287:391 -404.
[8]IJkel W F,van STRIEN E A,HELDENS J G,et al.Sequence and organization of the Spodoptera exigua multicapsid nucleopolyhedrovirus genome[J].J Gen Virol,1999,80(Pt 12):3289-3304.
[9]YANASE T,YASUNAGA C,HARA T,et al.Coinfection of Spodoptera exigua and Spodoptera frugiperda cell lines with the nuclear polyhedrosis viruses of Autographa californica and Spodoptera exigua [J].Intervirology,1998,41:244-252.
[10]O'REILLY D R,MILLER L K,LUCKOW V A.Baculovirus Expression Vectors:A Laboratory Manual[M].New York:W H Freeman,1992:256-258.
[11]ISHIKAWA H,IKEDA M,YANAGIMOTO K,et al.Induction of apoptosis in an insect cell line,IPLBLd652Y,infected with nucleopolyhedroviruses[J].J Gen Virol,2003,84:705 -714.
[12]PEARSON M N,ROHRMANN G F.Transfer,incorporation,and substitution of envelope fusion proteins among members of the Baculoviridae,Orthomyxoviridae,and Metaviridae(insect retrovirus)families[J].J Virol,2002,76:5301 -5304.
[13]LACOUNT D J,FRIESEN P D.Role of early and late replication events in induction of apoptosis by baculoviruses[J].J Virol,1997,71:1530 -1537.
[14]PRIKHOD'KO E A,MILLER L K.Induction of apoptosis by baculovirus transactivator IE1 [J].J Virol,1996,70:7116-7124.
[15]THIEM S M,MILLER L K.Identification,sequence,and transcriptional mapping of the major capsid protein gene of the baculovirus Autographa californica nuclear polyhedrosis virus [J].J Virol,1989,63:2008 -2018.
