抚育对不同坡向“栽针保阔”红松林群落结构的影响
2011-01-17于晓梅屈红军
于晓梅 屈红军
(东北林业大学,哈尔滨,150040)
抚育对不同坡向“栽针保阔”红松林群落结构的影响
于晓梅 屈红军
(东北林业大学,哈尔滨,150040)
以长白山区“栽针保阔”红松林为研究对象,采用样带网格调查和重要值研究法,研究了不同抚育方式对阔叶红松林群落结构的影响效果。结果表明:随着透光抚育强度加大,不同坡向的“栽针保阔”红松林群落中,红松优势地位提升,阴坡和半阳坡应保持较低郁闭度,阳坡应保持中等郁闭度;半阳坡红松重要值提升幅度(0.300~0.708)>阳坡(0.271~0.516)>阴坡(0.119~0.470),通过“栽针保阔”途径恢复阔叶红松林要充分考虑坡向环境因子。
透光抚育;红松林;群落结构;栽针保阔;坡向
阔叶红松林是中国东北东部山区地带性顶极植被,具有林地生产力高、森林生态环境优良、生物多样性良好、稳定性强等优点[1]。但由于半个多世纪的过度利用与破坏,原始阔叶红松林所剩无几,仅在自然保护区有少量的分布,目前已退化为天然次生林和人工林。因此,有关阔叶红松林的恢复是科学界普遍关注的问题。一方面,在自然保护区开展原始红松林结构、功能、动态及经营等方面的研究[2-4],揭示了原始红松林群落的发生发展规律性,不仅为原始红松林的保育与可持续经营提供了科学依据,而且也为东北次生林和人工林的经营提供了可借鉴的参照系统。另一方面,在次生林区开展了次生林的林型分类、结构、功能、动态及经营等方面的系统研究[5-7],探讨了红松混交林有效恢复与经营途径,并提出了适合东北森林特点的“栽针保阔动态经营体系”[8-10],以期通过在次生林林隙栽植红松等针叶树种,并在次生林不同发展阶段实施保阔、引阔和选阔,人工促进次生林向红松混交林方向的演替进程,为东北次生林的经营指明了发展方向。但目前“栽针保阔”恢复途径中,林下栽植的红松幼树生长缓慢,郁闭成林较晚等问题,给阔叶红松林的恢复和发展带来了一定困难,这是由于“栽针保阔”思想发展时间相对较短,其实践效果还需时间检验。因此,需进一步深入系统地长期定位研究,不断充实和完善该理论体系。本文对长白山林区不同坡向、不同抚育强度的“栽针保阔”红松林群落结构(重要值)进行研究,探讨不同抚育强度对“栽针保阔”红松林群落结构的影响,以期为长白山林区经营及阔叶红松林资源的恢复提供科学依据。
1 研究地区概况
研究地点位于长白山林区白河林业局春雷林场(41°43′~42°26′N,127°42′~128°17′E)。本区属受季风影响的温带大陆性气候,全年平均气温为3.9℃左右,年降水量600~700mm,土壤为发育在火山灰沙砾土上的暗棕色森林土。本区植物区系属于长白山植物区系,地带性植被以红松为主的红松阔叶混交林。
2 研究方法
2.1 样地设置及调查
研究区位于春雷林场14林班(1979年林冠下栽植红松)和16林班(1993年林冠下栽植红松),前者位于东南坡(半阳坡)和南坡(阳坡),后者位于东北坡(阴坡)。试验于1996年开始,分别在1979年与1993年林冠下栽植红松的林分中设置不同上层透光抚育强度(郁闭度:0(皆伐)、0.2、0.4、0.6、0.8(对照))处理试验(各处理水平面积均为1 hm2)。本调查标准地设置的方法:在不同时期及不同上层透光抚育强度处理试验区各设置3块30m×20m的标准地,共设置45块标准地。采用标准地样带网格调查方法,将各标准地均匀划分为24个5m×5m的小样方,对样地内乔木进行每木调查,测定乔木的种类、数量、频度、胸径及树高等,研究群落的树种组成结构。
2.2 重要值分析方法
依据群落各组成树种的重要值,判定各树种在试验群落重点地位及作用,分析不同抚育强度对群落树种组成结构的影响。
重要值=(相对密度+相对频度+相对盖度)/3。其中:相对密度=某个种的株数/所有种的总株数;相对频度=某个种在样方中出现的次数/所有种出现的总次数;相对盖度=某个种的胸高断面积/所有种的胸高断面积之和。
3 结果与分析
阴坡、半阳坡、阳坡“栽针保阔”红松林群落的树种组成结构与透光强度有着密切的关系,不同透光强度的群落树种组成结构存在明显的差异性。
3.1 阴坡“栽针保阔”红松林群落结构
未透光群落结构:未透光抚育(郁闭度0.8)的“栽针保阔”红松林群落,以阔叶树种占绝对优势地位(见表1)。其群落由8个树种组成,重要值从大到小排序为紫椴(Tilia amurensis)、春榆(Ulmus davidiana)、蒙古栎(Quercus mongolica)、红松(Pinus koraiensise)、黄波椤(Phellodendron amurensis)、枫桦(Betula costata)、色木槭(Acer mono)和白牛槭(A.mandshuricum),形成了以紫椴为优势种,春榆、蒙古栎和红松为亚优势种,黄波椤、枫桦、色木槭和白牛槭为伴生种的红松阔叶混交林群落。
半透光群落结构:半透光抚育(郁闭度0.2~0.6)的“栽针保阔”红松林群落,均以红松占绝对优势地位(见表1)。郁闭度0.6群落,由10个树种组成,重要值从大到小排序为红松、胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、黄波椤、山杨(Populus davidiana)、紫椴、山梨(Pyrus ussuriensis maxim)、旱柳(Salix matsudanna)、色木槭和山槐(Maackia a-murensis),形成了以红松和胡桃楸为优势种,水曲柳和黄波椤为亚优势种,山杨、紫椴、山梨、旱柳、色木槭和山槐为伴生种的红松林群落。郁闭度0.4群落,乔木树种由11个树种组成,重要值从大到小排序为红松、胡桃楸、春榆、水曲柳、山杨、山槐、色木槭、黄波椤、紫椴、山梨和旱柳,形成以红松为优势种,胡桃楸、春榆和水曲柳为亚优势种,山杨、山槐、色木槭、黄波椤、紫椴、山梨和旱柳为伴生种的红松林群落。郁闭度0.2群落,乔木树种由13个树种组成,重要值从大到小排序为山杨、红松、白桦(Betula platyphylla)、山槐、蒙古栎、黄波椤、春榆、山梨、紫椴、胡桃楸、色木槭、白牛槭和沙松(Abies holophylla Maxim.),形成了以山杨和红松为优势种,白桦、山槐、蒙古栎、黄波椤、春榆、山梨、紫椴、胡桃楸、色木槭、白扭槭和沙松为伴生种的红松林群落。
全透光群落结构:全透光抚育(郁闭度0)的“栽针保阔”红松林群落,也是以红松占绝对优势地位(见表1)。群落由7个树种组成,重要值从大到小排序为红松、黄波椤、春榆、胡桃楸、紫椴、白桦和蒙古栎,形成了以红松为优势种,黄波椤、春榆和胡桃楸为亚优势种,紫椴、白桦和蒙古栎为伴生种的红松林群落。
由此可见,随着抚育强度的加大,阴坡红松在群落中的地位得到提升,而主要阔叶树种紫椴和蒙古栎在群落中地位却逐渐下降,使得红松逐步取代了原次生林群落优势树种紫椴而成为群落的优势种。水曲柳、黄波椤和胡桃楸取代了蒙古栎的亚优势种的地位上升为亚优势种,春榆的亚优势种地位基本保持不变。透光抚育使群落伴生种数量均有所增加(增加了3~7种,除皆伐少1种外)。此外,透光抚育尽管均使红松在群落中的地位有所提升,但不同抚育强度的影响效果存在差异性。郁闭度在0.2~0.6时,透光抚育对红松的影响程度相近(0.303~0.325),而皆伐则有较大幅度提高(0.470)。因此,采取“栽针保阔”途径恢复红松林群落初期,透光抚育强度应保持较低郁闭度。
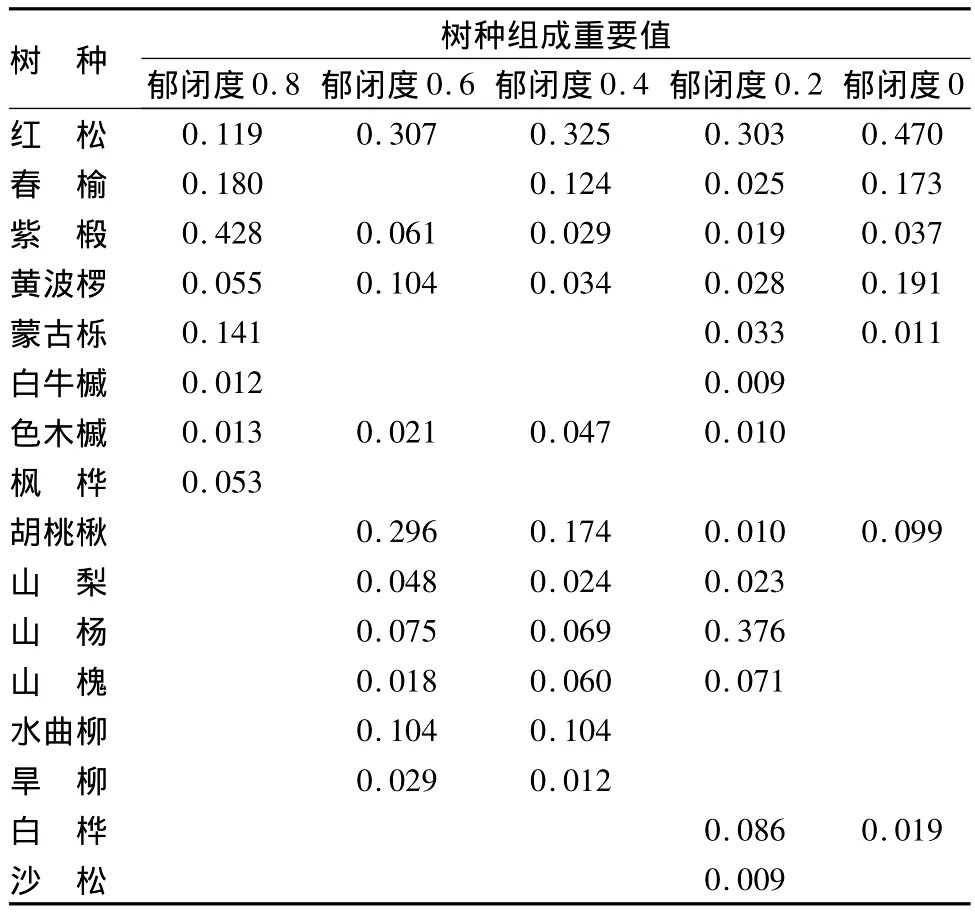
表1 阴坡“栽针保阔”红松林群落树种组成(重要值)
3.2 半阳坡“栽针保阔”红松林群落结构
未透光群落结构:未透光抚育(郁闭度0.8)的“栽针保阔”红松林群落,以阔叶树种占优势地位(见表2)。其群落由8个树种组成,重要值从大到小排序为紫椴、红松、蒙古栎、春榆、山杨、山槐、白牛槭和色木槭,形成了以紫椴和红松为共优势种,蒙古栎为亚优势种,春榆、山杨、山槐、白牛槭和色木槭为伴生种的红松林群落。

表2 半阳坡“栽针保阔”红松林群落树种组成(重要值)
半透光群落结构:半透光抚育(郁闭度0.2~0.6)的“栽针保阔”红松林群落,均以红松占绝对优势地位(见表2)。郁闭度0.6群落,由8个树种组成,重要值从大到小排序为紫椴、红松、蒙古栎、山槐、山杨、黄波椤、春榆和假色槭(Acer pseudo-sieboldianum(Pax.)Komarov),形成了以紫椴和红松为共优势种,蒙古栎为亚优势种,山槐、山杨、黄波椤、春榆和假色槭为伴生种的红松林群落。郁闭度0.4群落,由7个树种组成,重要值从大到小排序为红松、紫椴、蒙古栎、春榆、黄波椤、山槐和山梨,形成了以红松为优势种,紫椴和蒙古栎为亚优势种,春榆、黄波椤、山槐和山梨为伴生种的红松林群落。郁闭度0.2群落,由6个树种组成,重要值从大到小排序为红松、春榆、紫椴、蒙古栎、花曲柳(Fraxinus rhynchophylla)和山杨,形成了以红松为优势种,春榆和紫椴为亚优势种,蒙古栎、花曲柳和山杨为伴生种的红松林群落。
全透光群落结构:全透光(郁闭度0)的“栽针保阔”红松林群落,以红松占绝对优势地位(见表2)。其群落由4个树种组成,重要值从大到小排序为红松、春榆、紫椴和胡桃楸,形成了以红松为优势种,春榆为亚优势种,紫椴和胡桃楸为伴生种的红松林群落。
由此可见,随着透光抚育强度的加大,半阳坡红松在群落中的地位得到不断地提升,进而逐步成为群落的优势种,原次生林群落优势种紫椴的地位不断下降,逐步降为亚优势种和伴生种;原次生林群落的亚优势种蒙古栎地位基本保持不变,而春榆则由伴生种逐步上升为亚优势种。此外,群落组成树种数略有减少(皆伐下降4种,其他下降为1~2种)。因此,采取“栽针保阔”途径恢复红松林群落中期,透光抚育强度也应保持较低郁闭度。
3.3 阳坡“栽针保阔”红松林群落结构
未透光群落结构:未透光抚育(郁闭度0.8)的“栽针保阔”红松林群落,以蒙古栎占优势地位(见表3)。其群落由6个树种组成,重要值从大到小排序为蒙古栎、红松、紫椴、春榆、黄波椤、枫桦和花曲柳,形成了以蒙古栎和红松为优势种,紫椴为亚优势种,春榆、黄波椤、枫桦和花曲柳为伴生种的红松林群落。
半透光群落结构:半透光抚育(郁闭度0.2~0.6)的“栽针保阔”红松林群落,均以红松占优势地位(见表3)。郁闭度0.6群落,由6个树种组成,重要值从大到小排序为红松、蒙古栎、紫椴、春榆、假色槭和山槐,形成了以红松和蒙古栎为优势种,紫椴为亚优势种,春榆、假色槭和山槐为伴生种的红松林群落。郁闭度0.4群落,乔木层由7个树种组成,重要值从大到小排序为红松、蒙古栎、紫椴、春榆、花曲柳、枫桦和黄波椤,形成了以红松为优势种,蒙古栎和紫椴为亚优势种,春榆、花曲柳、枫桦和黄波椤为伴生种的红松林群落。郁闭度0.2群落,由6个树种组成,重要值从大到小排序为红松、紫椴、春榆、蒙古栎、山杨和山槐,形成了以红松为优势种,紫椴、春榆和蒙古栎为亚优势种,山杨和山槐为伴生种的红松林群落。

表3 阳坡“栽针保阔”红松林群落树种组成(重要值)
全透光群落结构:全透光(郁闭度0)的“栽针保阔”红松林群落,以红松占绝对优势地位(见表3)。其群落由5个树种组成,重要值从大到小排序为红松、紫椴、春榆、山杨和蒙古栎,形成了以红松为优势种,紫椴、春榆和山杨为亚优势种,蒙古栎为伴生种的红松林群落。
由此可见,随着透光抚育强度的加大,阳坡红松在各群落中的地位得到不断提升,逐步成为群落的优势种,原次生林群落的优势种蒙古栎的地位不断下降,逐步降为亚优势种和伴生种。紫椴和春榆则由伴生种逐步上升为亚优势种。红松的重要值表现为中等郁闭度条件下最大(0.401~0.516)。因此,在阳坡采取“栽针保阔”途径恢复红松林群落的中期,透光抚育强度应保持中等郁闭度。
4 结论与讨论
阴坡“栽针保阔”红松林群落,在采取不同强度的透光抚育后,均形成了以红松为优势种的针阔叶混交林群落,但各群落的树种组成结构却有所不同。对照群落,主要由紫椴、春榆、蒙古栎和红松组成针阔混交林。随着透光抚育强度的加大,红松在群落中的地位得到提升而成为优势种,形成了红松与胡桃楸、黄波椤、水曲柳、春榆为主的针阔混交林。
半阳坡“栽针保阔”红松林群落,在采取不同强度的透光抚育后,均形成了以红松为优势种的针阔叶混交林群落,但各群落的树种组成结构却有所不同。对照群落,主要由紫椴、红松和蒙古栎组成针阔混交林。随着透光抚育强度的加大,红松在群落中的优势地位进一步得到提高,紫椴优势地位下降,形成了以红松与春榆、蒙古栎、紫椴为主针阔混交林。
阳坡“栽针保阔”红松林群落,在采取不同强度的透光抚育后,均形成了以红松为优势种的针阔叶混交林群落。对照群落,主要由蒙古栎、红松和紫椴形成的针阔混交林。随着抚育强度的加大,红松在群落中的优势地位提升,蒙古栎优势地位下降,形成了以红松与蒙古栎、紫椴和春榆为主的针阔混交林。
采取不同的透光抚育后,红松在群落中的地位都得到提升,但在不同坡向其影响效果有所差异。半阳坡红松重要值(0.300~0.708)提升幅度大于阳坡(0.271~0.516),主要是由于在阳坡立地上,阔叶树种提高的幅度也比较大,对红松的竞争抑制相对较强。半阳坡红松重要值提高的幅度更大于阴坡(0.119~0.470),这说明“栽针保阔”途径下的红松生长也需光照条件,“幼年耐阴”的特性不应成为其长期在庇荫条件下生长的依据。因此,通过“栽针保阔”这条有效途径恢复阔叶红松林,要充分考虑坡向环境因子,特别是在半阳坡,逐步加大伐除阔叶树的强度,更有利于红松的生长。
面对东北林区红松林资源消耗殆尽的局面,目前就红松的生长而言,提早结实或分叉与否并不重要,重要的是如何使更新的红松尽快生长,增加地带性顶极植被的本底,为恢复东北的阔叶红松林创造有利的群落环境条件。
[1] 代力民.红松混交林生态与经营实践[M].北京:科学出版社,2002.
[2] 詹鸿振,徐贵林,刘传照,等.凉水自然保护区阔叶红松林的群落结构初报[J].东北林学院学报,1984,12(增刊):8-20.
[3] 郑景明,罗菊春.长白山阔叶红松林结构多样性的初步研究[J].生物多样性,2003,11(4):295-302.
[4] 郝占庆,陶大立,赵士洞.长白山北坡阔叶红松林及其次生白桦林高等植物物种多样性比较[J].应用生态学报,1994,5(1):16-23.
[5] 陈大珂,周晓峰,祝宁,等.天然次生林:结构·功能·动态与经营[M].哈尔滨:东北林业大学出版社,1994.
[6] 李新彬,王襄平,罗菊春,等.干扰对长白山北坡次生林群落结构和树种多样性的影响[J].林业科学,2006,42(2):105-110.
[7] 沈海龙,张群,范少辉,等.次生林群落结构特性对红松幼树生长的影响[J].林业科学研究,2004,17(5):610-615.
[8] 周晓峰.红松阔叶林的恢复途径:栽针保阔[J].东北林学院学报,1982,10(增刊):18-28.
[9] 陈大珂,周晓峰,丁宝永,等.黑龙江省天然次生林研究(Ⅰ):栽针保阔的经营途径[J].东北林学院学报,1984,12(4):1-11.
[10] 陈大珂,周晓峰,丁宝永,等.黑龙江省天然次生林研究(Ⅱ):动态经营体系[J].东北林学院学报,1985,13(1):1-18.
Effect of Tending on Community Structure of Broad-Leaved Korean Pine Forests at Different Slope Aspects
/Yu Xiaomei,Qu Hongjun(School of Forestry,Northeast Forestry University,Harbin 150040,P.R.China)//Journal of Northeast Forestry University.-2011,39(6).-10~12
Tending;Korean pine forests;Community structure;Planting conifers and protecting broadleaves;Slope aspects
S753.51+1
于晓梅,女,1973年5月生,东北林业大学图书馆,馆员。E-mail:yuxiaomei1973@hotmail.com。
屈红军,东北林业大学图书馆,副研究馆员。E-mail:quhongjun2001@yahoo.com.cn。
2010年10月12日。
责任编辑:张 玉。
A survey was conducted to study the effect of different tending modes on the community structure of broad-leaved Korean pine forests after planting conifers and protecting broadleaves in terms of species importance value by the grid method.Result show that the Korean pine in the broad-leaved forests gradually became dominant with increasing tending intensity.The north-facing slope and semi-south slope should retain a lower canopy density,while the south-facing slope should retain a moderate canopy density.The increase extent of importance value for Korean pine in semi-south slope ranged from 0.300-0.708,followed by south-facing slope(0.271-0.516)and north-facing slope(0.119-0.470).It shows that more attention should be paid to slope aspect during the recovery process of Korean pine forests after planting conifers and protecting broadleaves.
