山东半岛南部近岸海域渔业资源群落结构的季节变化*
2011-01-10张秀梅张沛东黄国强
李 涛,张秀梅,张沛东,黄国强
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
山东半岛南部濒临黄海,是多种海洋生物的产卵场、索饵场和传统渔场[1]。作为我国重要的渔业作业海域,该海区蕴藏丰富的自然资源,在满足市场需求,出口创汇和稳定渔业人口生活等方面发挥了重要作用[2]。近年来,由于过度捕捞和环境变化等多种因素的影响,渔业资源衰退,导致渔获物中传统经济种类比例下降,一些低质种类成为主要捕捞对象[3]。
近岸海域的突出特点之一是有众多河流入海,受人类活动影响较大,近岸海域和沉积物中的营养盐与有机碎屑都比近海和外海含量要高。因此,该区初级生产力也比较高[4]。近年来,关于黄海海域浮游动植物、水文环境、生物多样性等可见相关调查与报道[5-8]。但对山东半岛南部近岸海域渔业资源的大面调查尚未开展。本研究借助我国近海海洋综合调查与评价专项(SD-908)的调查工作,在山东半岛南部近岸海域开展了4个季度的底拖网调查,根据渔获物取样结果,对该区域渔业生物资源种类组成、群落结构特征等进行了分析,以为该海域的渔业管理和资源的科学利用提供基础依据。
1 材料与方法
本调查租用鲁乳渔4695、鲁乳渔4696双拖捕捞作业船只。2艘船长均为19.6 m、功率79.4 kw、最大航速10 nmile。作业网具为有翼单囊拖网,网具主尺寸113.15 m×83.99 m,身网网目(起)820目,身网网目(终)660目,身网节数241节。拖网调查范围为山东半岛南部119°33′E~122°0′E,35°19′N~36°40′N沿岸海域。4个航次调查时间为2006年7月下旬、12月底,2007年4月上旬、11月下旬,依据908合同要求共设置15个站位,站位具体位置参照图1,每站拖网时间1 h,拖速为3 nmile/h。胶州湾内两站位QD-09,QD-15由于养殖区的限制,实际拖网位置相距很近,在图中显示近似重合。
利用Margalef的种类丰富度指数R、Shannon-W ener多样性指数H′、Pielou均匀度指数J′来分析渔业资源群落生态多样性[9-10]。由于不同种类及同种类个体大小差异很大,Wilhm提出用生物量表示的多样性更接近种类间能量分布,因此文中根据生物量计算群落种类多样性[10-12]。
Margalef的种类丰富度指数R:R=(S-1)/LnW Shannon-Wiener多样性指数Pielou均匀度指数J′:J′=H′/LnS
以上式中,S为种类数,W为总渔获量(kg),Pi为i种鱼占总渔获重量的比例。
根据相对生物量(kg/h)平方根转换计算的B ray-Curtis相似性系数矩阵,应用Cluster聚类分析来研究群落结构[13]。单因子相似性分析(ANOSIM)用于不同组群落结构差异显著性检验[14]。相似性百分比分析(SIM PER)用于分析造成各组内群落结构相似的典型种以及造成不同组之间群落结构差异的分歧种[13]。
2 结果
2.1 渔业资源结构
4个季度调查共捕获渔获种类72种,其中中上层鱼类8种,底层鱼类39种,甲壳类19种,头足类6种,分别占总种数的11.1%、54.2%、26.4%和8.3%。
夏季航次渔获种类中,鱼类的平均网获量为12.254 kg/h,甲壳类为4.54 kg/h,头足类为1.34 kg/h。
冬季航次渔获种类中,鱼类的平均网获量为1.854 kg/h,甲壳类为0.21 kg/h,头足类为4.62 kg/h。
春季航次渔获种类中,鱼类的平均网获量为0.671 kg/h,甲壳类为0.345 kg/h,头足类为0.278 kg/h。
秋季航次渔获种类中,鱼类的平均网获量为4.272 kg/h,甲壳类为3.590 kg/h,头足类为2.425 kg/h。
表1为山东半岛南部近岸海域4个季度渔获物重量和尾数组成。夏季底层鱼类和甲壳类占总渔获量的比例较高,分别为46.1%和25.0%;底层鱼类占总渔获尾数的45.0%。冬季底层鱼类和头足类占总渔获量的比例较高,为69.1%。春、秋2季各生态类渔获重量相对平均,春季底层鱼类渔获量占总渔获量比例略高,为37.4%。秋季则是甲壳类渔获量占总渔获量比例略高,为34.9%。

表1 山东半岛南部近岸海域渔获物组成Table 1 The composition of fishery resources in the coastal waters of Southern Shandong Peninsular
2.2 优势种组成
表2为4个季度调查中渔获重量列前5位,且出现在较多站位的优势种组成。夏季调查以鹰爪虾(Trachypenaeus curvirostris)和口虾蛄(Oratosquilla oratoria)优势度较高,分别占总渔获重量的14.5%和10.1%,5个种类合计占总渔获重量的46.4%,占总个体数的49.5%。冬季调查以尖海龙(Syngnathusa acus)和赤鼻棱鳀(Thryssa kamm alensis)占较大优势,分别占总渔获量的11.5%和7.8%,5个种类合计占总重量的30.9%,占总个体数的37.2%。春季调查以口虾蛄和方氏云鳚(Enedrias fangi)占较大优势,分别占总渔获量的16.2%和13.1%,5个种类合计占总重量的52.5%,占总个体数的70.1%。秋季调查以剑尖枪乌贼(Loligo edulis)和口虾蛄占较大优势,分别占总渔获量的20.1%和12.2%,5个种类合计占总重量的58.1%,占总个体数的93.9%。

表2 山东半岛南部近岸海域四季渔获量前5位种类的组成Table 2 Top 5 species by weight in the coastal watersof Southern Shandong Peninsular during four seasons
2.3 多样性特征
夏季渔业生物群落多样性指数在青岛沿岸海域的ZD-QD256、ZD-QD252、ZD-QD239几个站位较高。如站位ZD-QD256,种类丰度指数R、多样性指数H′和均匀度指数J′分别为3.459、0.722和2.126,明显高于其他站位,如丁字湾以东乳山近海的站位JC-HH 097(种类丰度指数R、多样性指数H′和均匀度指数J′分别为1.861、0.479和1.191)、胶南古镇口以西的近海站位JC-HH148(种类丰度指数R、多样性指数H′和均匀度指数J′分别为1.833、0.690和1.822)等站位。其他3个季节,多样性指数同样呈现类似的高低区别,即丁字湾以东和古镇口以西的站位在多样性上低于青岛沿岸的站位。山东半岛南部近岸海域4个季节渔业资源多样性指数如表3所示。
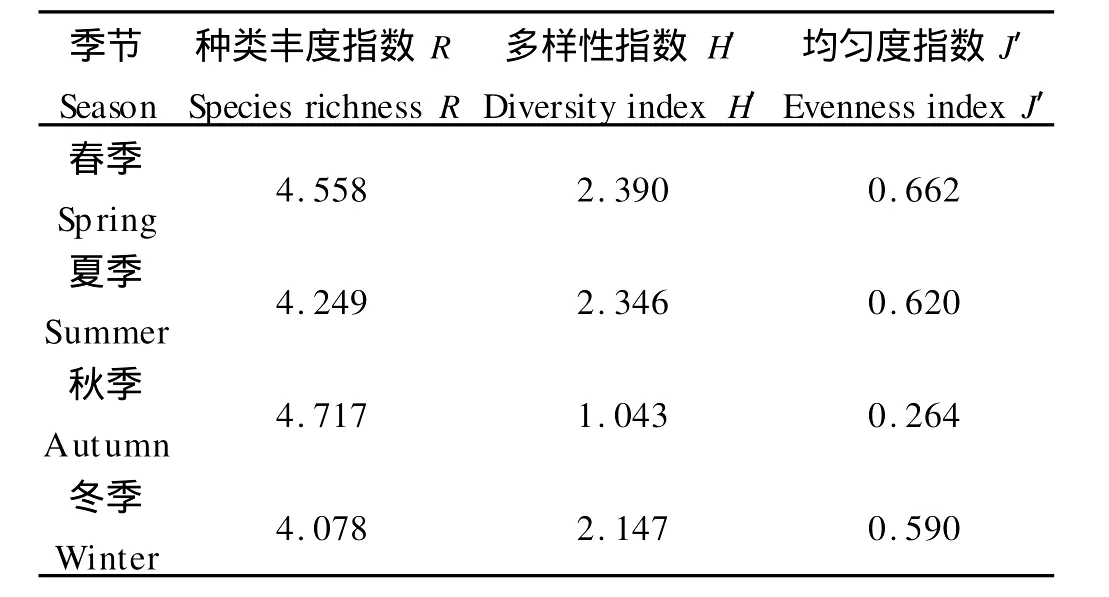
表3 山东半岛南部近岸海域渔业资源多样性指数Table 3 Diversity index of fishery resources in the coastal waters of Southern Shandong Peninsular
2.4 群落结构分析
通过Cluster聚类分析,夏季渔业资源群落分为3组,组1包括3个站位,其中ZD-QD249、ZD-QD252是处于调查海域南部,相对陆地较远的站位;组2包括8个站位,主要由ZD-QD213、ZD-QD228、ZD-QD239、ZD-QD245以及ZD-QD235等距岸线适中的站位组成;组3包括QD-09、QD-15、JC-HH097和ZD-QD256 4个站位,是距岸线较近以及位于胶州湾内的站位组成的聚类组。
冬季,渔业资源群落也大致可分为3组,由于JCHH 097和ZD-QD213站位的捕捞个体过少,所以不予分组。ZD-QD256、ZD-QD252和JC-HH148 3个站位归为组1,是调查海域南部的3个站位;ZD-QD228、ZD-QD239、ZD-QD235、ZD-QD237、ZD-QD226和ZDQD245 6个站位归为组2,主要位于青岛沿岸海区,并且是距岸线相对较远的几个站位;另外4个站位除ZDQD249相对陆地较远外,QD-09、QD-15、ZD-QD218都是属于青岛近海及胶州湾内的近岸站位,在聚类分析上呈现较高的相似性,归为组3。
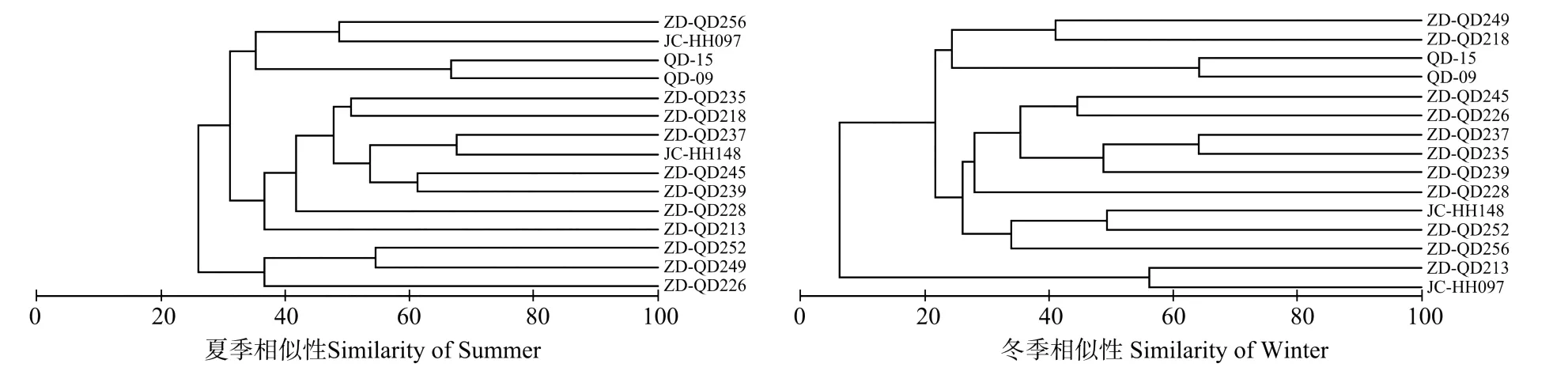
图2 山东半岛南部近岸海域夏季与冬季渔业资源群落聚类分析Fig.2 Cluster analysis for the fish resources assemblages based on the Bray-Curtis similarity matrices in the coastal watersof Southern Shandong Peninsular in summer and winter
春季,将山东半岛南部近岸海域渔业资源群落分为3组,由于ZD-QD239和ZD-QD249 2个站位的捕捞个体过少,所以不予分组。ZD-QD213、JC-HH097和QD-15 3个站位归为组1,其中ZD-QD213、JC-HH 097是调查最北部的2个站位;ZD-QD256和ZD-QD245 2个站位归为组2,位于胶州湾以西,且2站位距离较近;另外8个站位包括ZD-QD226、ZD-QD228、ZDQD235、ZD-QD218、ZD-QD237以及QD-09等都是位于青岛近岸的站位,在聚类分析上呈现较高的相似性,归为组3。
秋季,同样将该海域渔业资源群落分为3组,ZDQD218、ZD-QD249、ZD-QD235和ZD-QD252 4个站位归为组1,主要是调查中距岸线较远的站位;ZDQD239、ZD-QD237、JC-HH097、ZD-QD226、ZDQD213等9个站位归为组2,主要位于调查海域的北部和中部,并包含胶州湾内的2个站位QD-09及QD-15;另外2个站位ZD-QD228和JC-HH 148,在聚类分析上呈现较高的相似性,归为组3。
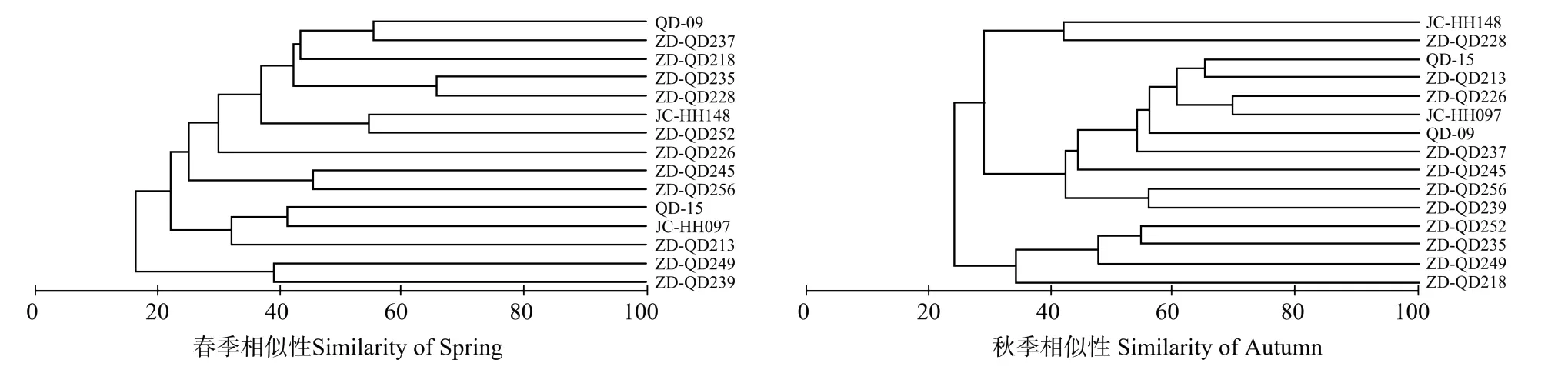
图3 山东半岛南部近岸海域春季与秋季渔业资源群落聚类分析Fig.3 Cluster analysis for the fish resources assemblages based on the Bray-Curtis similarity matrices in the coastal waters of Southern Shandong Peninsular in spring and autumn
ANOSIM分析表明,夏季不同站位组间群落结构种类组成差异极显著(R=0.762,P<0.001),站位组两两之间差异显著(P<0.05)。冬季不同站位组间群落种类组成差异显著(R=0.633,P<0.001),站位组两两之间差异显著(P<0.05)。春季不同站位组间群落种类组成差异显著(R=0.701,P<0.001),除组1与组2之外,站位组两两之间差异显著(P<0.05)。秋季不同站位组间群落种类组成差异显著(R=0.865,P<0.001),除组1与组3之外,站位组两两之间差异显著(P<0.05)。
夏季组1~3各站位组内平均相似性分别为22.13%、35.50%和31.54%。表4列出了夏季各种生物对组内平均相似性贡献占4%以上的种类及其贡献百分比。组1的主要典型种包括尖海龙、日本鳀(Engrau lis japonicus)和鹰爪虾,以上3种对组内平均相似性的贡献达到91.7%,组2的主要典型种包括鹰爪虾、口虾蛄、剑尖枪乌贼、日本蟳(Charybdis japonica)和细条天竺鱼(A pogon lineatus),以上5种对组内平均相似性的贡献达到90.55%,组3的主要典型种包括鹰爪虾、剑尖枪乌贼、日本蟳、赤鼻棱鳀和中国明对虾(Fenneropenaus chinensis),以上5种对组内平均相似性的贡献达到92.52%。组1和组2;组1和组3;组2和组3之间的平均相异性分别为87.07%、90.30%和84.29%。表3亦列出了夏季各种生物对组间平均相异性贡献4%以上的种类及其贡献百分比。对不同组间平均相异性贡献较高的分歧种包括尖海龙、鹰爪虾、口虾蛄、日本鳀、剑尖枪乌贼等。

表4 夏季组内典型种对组内平均相似性、组间分歧种对组间平均相异性的贡献百分比Table 4 Typifying species for different groups and their contributions to the average within-group similarity,discriminating species and their contributions to the average between-group dissimilarity during summer in the coastal waters of Southern Shandong Peninsular(≥4%)
冬季组1~3各站位组内平均相似性分别为19.28%、25.64%和16.97%。表5为冬季各种生物对组内平均相似性贡献占4%以上的种类及其贡献百分比。组1的主要典型种包括尖海龙、赤鼻棱鳀和剑尖枪乌贼,以上3种对组内平均相似性的贡献达到91.51%,组2的主要典型种包括尖海龙、剑尖枪乌贼、双喙耳乌贼(Sepiola birostrat)、鲜明鼓虾(A lpheus d istinguendus)、三疣梭子蟹(Portunus trituberculatus)、中国毛虾(Acetes chinensis)和赤鼻棱鳀,以上几种对组内平均相似性的贡献达到86.36%,组3的主要典型种包括梭鱼(Chelon haem atocheilus)、双喙耳乌贼和细螯虾(Leptochela gracilis),以上3种对组内平均相似性的贡献达到89.05%。组1和组2;组1和组3;组2和组3之间的平均相异性分别为89.58%、95.56%和93.09%。表4亦列出了冬季组间平均相异性贡献4%以上的种类及其贡献百分比。对不同组间平均相异性贡献较高的分歧种包括赤鼻棱鳀、尖海龙、中国毛虾、剑尖枪乌贼、细螯虾、梭鱼和双喙耳乌贼等。

表5 冬季组内典型种对组内平均相似性、组间分歧种对组间平均相异性的贡献百分比Table 5 Typifying species for different groups and their contributions to the average within-group similarity,and discriminating species and their contributions to the average between-group dissimilarity during w inter in the coastal waters of Southern Shandong Peninsular(≥4%)
春季组1~3各站位组内平均相似性分别为24.24%、30.00%和27.96%。表6为春季各种生物对组内平均相似性贡献占4%以上的种类及其贡献百分比。组1的典型种包括方氏云鳚、日本枪乌贼(Loligo japonicas)和口虾蛄,以上3种构成了对组内平均相似性的全部贡献,组2的主要典型种包括中颌棱鳀(Thryssam ystax)、玉筋鱼(Ammod ytes personatus)、绯(Callianym us richardsoni)、六丝矛尾鰕虎鱼(Amblychaeturichthys hexanem)和褐菖鲉(Sebastiscusm armoratus),以上5种对组内平均相似性的贡献达到90.47%,组3的主要典型种包括方氏云鳚、双喙耳乌贼和双斑蟳,以上3种对组内平均相似性的贡献达到81.74%。组1和组2;组1和组3;组2和组3之间的平均相异性分别为88.10%、88.55%和83.75%。表5亦列出了春季组间平均相异性贡献4%以上的种类及其贡献百分比。对不同组间平均相异性贡献较高的分歧种包括方氏云鳚、口虾蛄、中颌棱鳀、绯、玉筋鱼、日本枪乌贼、黄鲫(Setipinna taty)、赤鼻棱鳀、双喙耳乌贼、双斑蟳(Charybdis bim aculata)和葛氏长臂虾(Palaemon gravieri)等。
秋季组1~3各站位组内平均相似性分别为28.90%、42.25%和26.19%。表7为秋季各种生物对组内平均相似性贡献占4%以上的种类及其贡献百分比。组1的主要典型种为细巧仿对虾(Parapenaeopsis tenella),对组内相似性贡献高达94.79%,组2的主要典型种包括鹰爪虾、剑尖枪乌贼、口虾蛄和六丝矛尾鰕虎鱼,以上4种对组内平均相似性的贡献达到89.90%,组3的主要典型种包括鹰爪虾、剑尖枪乌贼、口虾蛄和双斑蟳,以上4种对组内平均相似性的贡献达到90.91%。组1和组2;组1和组3;组2和组3之间的平均相异性分别为92.56%、97.16%和87.04%。表6亦列出了秋季组间平均相异性贡献4%以上的种类及其贡献百分比。对不同组间平均相异性贡献较高的分歧种包括细巧仿对虾、鹰爪虾、剑尖枪乌贼、口虾蛄和六丝矛尾鰕虎鱼等。
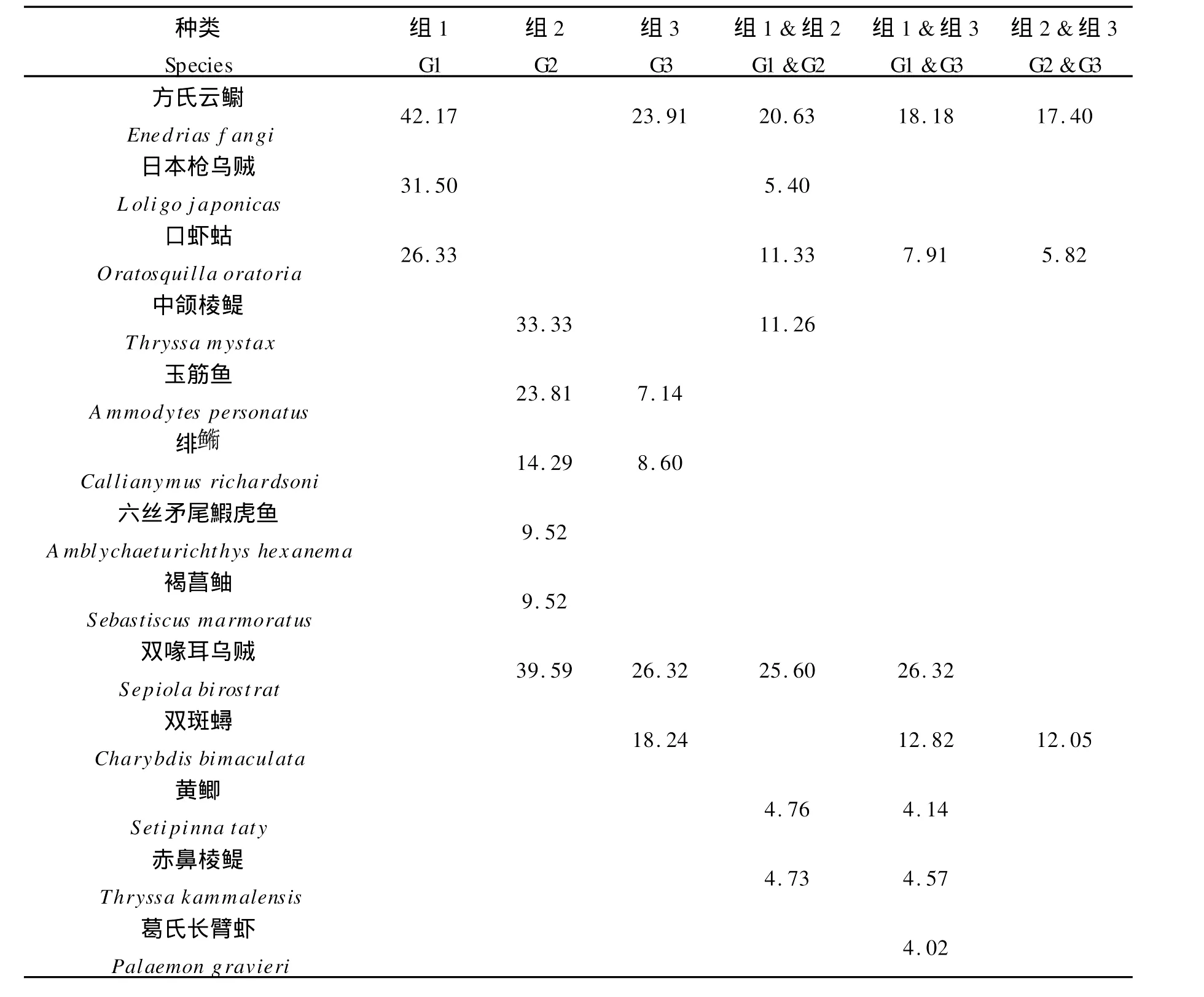
表6 春季组内典型种对组内平均相似性及组间分歧种对组间平均相异性的贡献百分比Table 6 Typifying species for different groups and their contributions to the average within-group similarity,and discriminating species and their contributions to the average between-group dissimilarity during sp ring in the coastal waters of Southern Shandong Peninsular(≥4%)

表7 秋季组内典型种对组内平均相似性、组间分歧种对组间平均相异性的贡献百分比Table 7 Typifying species for different groups and their contributions to the average within-group similarity,and discriminating species and their contributions to the average between-group dissimilarity during autumn in the coastal waters of Southern Shandong Peninsular(≥4%)
3 讨论
3.1 种类组成
山东近海地处暖温带季风气候区,每年洄游于渤、黄海或东海之间的许多游泳生物都在山东半岛近岸海域过路、产卵或索饵,所以山东半岛近岸海域既是渔业生物资源的重要产卵场和索饵场,同时又是生物资源进出渤海的重要通道[4]。本次调查的海域,生物资源基本可划分为2个生态类群,即地方性和洄游性资源。地方性资源主要栖息在河口、岛礁和浅水区,随着水温的变化,作季节性深-浅水生殖、索饵和越冬移动,迁移距离较短,洄游路线不明显,如多种暖温性地方种类;而洄游性渔业资源,主要为暖温性和暖水性种类,分布范围广、洄游距离长,为主要渔业种类[15]。根据历史调查,山东半岛南部近海栖息鱼类有100余种以上[16-17],程济生等于1998年在山东半岛近岸海域的调查中共获鱼类63种,分隶11目,41科。曾晓起等根据2003年10月~2004年5月逐月采集胶州湾及其邻近海域作业的定置网和底拖网渔获物,共记述了106种渔业生物,包括鱼类58种和无脊椎动物48种[18]。本研究调查共计渔获种类72种,其中鱼类共47种,隶属9目,29科,43属。黄鮟鱇(Lophius litulon)、小黄鱼等暖温性种类占鱼类种数的46.8%,赤鼻棱鳀、黄鲫、鲬(Platycephalus indicus)等暖水性种类占鱼类种数的40.4%,玉筋鱼,方氏云鳚等冷温性鱼类占12.8%。各季节出现的鱼类与其生活习性密切相关,以山东半岛南部近岸海域作为产卵场的一些底层鱼类,如小黄鱼、鲬等产卵期在5~6月;一些中上层鱼类,如黄鲫、方氏云鳚等部分群体,在4月份到达近岸产卵繁殖,故在4月上旬的春季拖网调查中,方氏云鳚和黄鲫的平均网获重量排在第二位和第三位,而个体较大的底层鱼类出现频率极低,使得渔获总重量和渔获个体平均重量在4个航次中都呈现最低。在夏、秋季拖网渔获物中底层鱼类增多,如小黄鱼、星康吉鳗的渔获重量分别占夏季鱼类总重量的6.44%、3.88%,秋季分别为16.03%、2.48%,而在冬季,小黄鱼、星康吉鳗的渔获重量分别仅占鱼类总重量的4.46%、0.17%。黄海地方性冷温种,如高眼鲽(Cleisthenes herzensteini)等,洄游距离较短,但也会随着季节变化进行深水→浅水→深水的越冬、生殖索饵洄游[3]。本研究的冬季和春季航次调查,近岸调查站位水温较低,表层水温在8~11℃之间,但由于水深较浅,青岛近岸和丁字湾以东站位的水深仅为20 m左右,因此未捕获到黄海冷水团中的冷温种大头鳕(Gadus macrocephalus)。高眼鲽在冬季航次未出现,春季航次仅在青岛近岸站位QD-245捕获到3尾。
根据本文表2优势种分析结果,明显看出4个航次均以甲壳类,头足类等无脊椎动物和部分小型低质鱼类为优势种。如口虾蛄,除冬季航次外,在其他3个季节均排在优势种的前两位,甲壳类的主要经济种鹰爪虾在夏季渔获物中占总重量的15.60%,排在优势种的第一位,在秋季占渔获物总重量的9.28%,排优势种的第三位,但在其他两个季节的渔获物中未出现,这一点与鹰爪虾在黄海渔汛期为6~7月(夏汛)及10~11月(秋汛)是相一致的。三疣梭子蟹在4个航次均有出现,除春季仅捕获3只外,其他3个季节的出现频率均匀,分别占夏秋冬3个航次渔获重量的1.43%、1.80%和2.64%。
3.2 聚类特征
分析结果表明,山东半岛南部近岸海域渔业资源群落的空间分布规律与生态环境的变化,诸如离岸距离、河流入海、地理位置等一系列因素关系密切,因而群落结构呈现一定的空间异质性,而在各季节航次调查中又有不同体现。随着离岸距离的增加,调查站位的水深也出现较大变化,如胶州湾内的2个站位,以及几个近岸站位如JC-HH148等,水深在15 m左右,离岸距离稍远的站位,如ZD-QD252等,水深在25 m左右甚至更大。四季渔业生物群落均可分为3个站位组,随季节的变化,站位分组也有较大区别。夏季调查时间为7月下旬,聚类分组明显由离岸距离决定,各站位分为近岸、远岸和距岸线适中的3个站位组,根据历史水文记录,8月份整个海区表层盐度降至全年最低,夏季聚类情况应与降水丰富,地表径流大量入海引起的近岸海域生物化学指标形成一定梯度有关,且夏季底层水温随离岸距离的增加,降温幅度最大,这一点在夏季调查中也很明显,近岸站位表层水温都在25℃以上,胶州湾内的站位表层水温达到26℃,而距离陆地较远的站位,如ZD-QD252,表层水温只有24.2℃。故盐度与水温是造成以离岸距离为聚类分组的形成因素;冬季,降水及沿岸径流最少,近岸低盐水很弱,因此聚类与夏季明显不同,根据Cluster聚类分析,古镇口以西的3个站位单独成组,尖海龙、赤鼻棱鳀和剑尖枪乌贼是该聚类组的主要典型种,其余站位分为近岸和远岸2组,区分近岸和远岸的主要分歧种是赤鼻棱鳀、梭鱼和细螯虾,这几种都较多的出现在离岸较远的站位;春季,调查海区水文状况处于由冬季向夏季分布特征的过渡阶段,海水增温缓慢且水温不稳定,这一点在春季调查中也很明显,站位ZD-QD213的表层水温只有8℃,而与之相距最近的站位ZD-QD218的表层水温却有11℃。胶州湾以西的2个站位呈现出单独分组聚类特征,主要典型种包括中颌棱鳀、玉筋鱼和绯,青岛海域8个站位聚类明显,双喙耳乌贼、方氏云鳚和双斑蟳是该组最主要的3个典型种,其余2个位于丁字湾以东的站位及胶州湾内的1个站位,以方氏云鳚、日本枪乌贼和口虾蛄为典型种而区分于其他2个站位组;秋季调查海区水温开始逐渐下降,沿岸降温较外海快、北部较南部快,冷水团向深层龟缩且势力逐渐减弱,调查站位表层水温在11~15℃之间,除去以剑尖枪乌贼、双斑蟳、鹰爪虾和口虾蛄4种无脊椎动物为主要典型种而单独分组的两个站位外,秋季离岸较远的几个站位因典型种细巧仿对虾而单独分组,其他九个站位,从调查海域最东端的石岛渔场到青岛近岸海域,呈现出较高相似性,主要典型种是剑尖枪乌贼、鹰爪虾和口虾蛄等。相似性百分比分析(SIM PER)表明各站位组的典型种同时又是各站位组间一定程度的分歧种,如赤鼻棱鳀、口虾蛄、双斑蟳、剑尖枪乌贼、短蛸和方氏云鳚等既是各组的典型种,又是组间的分歧种。分析认为组间群落结构种类组成的差异性主要是由于这些种类数量分布的空间差异和拖网调查的随机性造成的,而多样性的构成与生物量的多寡并不构成必然联系。
3.3 渔业资源分析
本次调查发现,山东半岛南部近岸海域渔业资源量水平较低,2006年夏、冬,2007年春、秋4个航次单位网次渔获量分别为18.1、6.9、1.3、10.3 kg/h。春季和秋季是传统的渔业汛期,渔获量应该处在高位。但本研究春季调查在4月上旬,而秋季调查在11月下旬进行,春季近岸水温尚未升高,鱼类未至近岸;11月下旬近岸水温迅速降低,在近岸浅水区活动的鱼类已向深水区移动,从而导致春、秋两季渔获量较低。另外,秋季调查时距伏季休渔期结束,捕捞活动重新开始已经有较长时间,而春季更是经过秋冬高强度的捕捞作业,使春季渔获量更低,这也说明高强度的捕捞活动对渔业资源产生了极大影响。本研究调查海域的优势种类主要为赤鼻棱鳀、长蛇鲻、方氏云鳚以及口虾蛄、剑尖枪乌贼、鹰爪虾等无脊椎动物。夏季5个优势种类占总重量的46.4%,冬季5个优势种类合计占总重量的30.9%,春季5个优势种类合计占总重量的52.5%,而秋季5个优势种类合计占总重量的58.1%,调查海区缺乏对资源量起支配作用的大宗经济型优势种类。一些传统经济种类,如真鲷(Chrysophrysmajor)、褐牙鲆(Paralichthys olivaceus)未出现。带鱼(Trichiurus lep turus)、许氏平鲉(Sebastes schlegeli)和银鲳(Strom ateoides argenteus)等在调查中所占渔获量比例较小,只有银鲳在冬季渔获物中单网渔获重量排在第四位。秋季渔获物中,带鱼在丁字湾以东的2个站位渔获较多,但在青岛近海和古镇口以西12个站位仅捕获8尾,许氏平鲉虽然在4个季节均有出现,但4个季节仅共捕获41尾。根据金显仕对山东南部海域1986年、1998年和2000年底拖网调查分析,日本鳀均占据优势种的第一位[15],而本研究调查中,日本鳀只有夏季排在优势种的第五位,其他曾经的优势种如高眼鲽、玉筋鱼等均未进入4个季节前5位的优势种之列。渔获物组成分析表明,多年生鱼类的生物学规格很小,当年生鱼和一龄鱼占据绝大比例。据报道,随着近年来捕捞对象向一些次级经济鱼类的转移,使得如玉筋鱼,黄鲫等许多低值经济鱼类产量亦有大幅下降[3]。在本研究调查中,单网渔获重量超过1 kg的单种鱼类只有夏季的长蛇鲻和日本鳀。鱼类资源量的下降,在某种程度上降低了对甲壳类、贝类等的捕食压力和饵料竞争压力,将有助于甲壳类等幼体的成活,提升诸如对虾、梭子蟹等重要经济种类的产量,但一个完整的生物群落,依托于稳定的生态系统和相互制约的生物链,任何一个等级的衰退或环节的缺失对于整个生物群落的安全都是致命的,要维护渔业资源的稳定,就必须着眼于整个海洋生态系统的保护。
本研究调查发现,渔获个体重量组成以小型个体占绝对优势,夏季渔获个体平均重量为10.96 g,冬季渔获个体平均重量11.51 g,春季渔获个体平均重量7.27 g,除去细巧仿对虾的影响,秋季个体平均重量为12.23 g,与夏冬两季节相当。无论哪个季节,渔获物均以小型低质种类及当年生幼鱼为主。由于调查站位均处在近岸较浅水域,会对渔获物组成产生很大影响。但总体而言,调查海域渔业资源已经充分或过度利用,资源的营养级下降,生命周期长、经济价值高的渔业资源已经严重衰退,长期的过度捕捞造成小型化、性早熟等种群衰退特征明显。因此,开展渔业资源养护应充分考虑资源衰退留下的空缺生态位,以及渔业生物群落结构的空间异质性,开展不同生态位物种的增殖放流,以达到维护生态系统平衡,保持渔业资源可持续利用之目的。
致谢:感谢中国海洋大学徐宾铎老师对论文的大力帮助,在此表示衷心感谢!
[1] 刘效舜,吴敬南,韩光祖,等.黄渤海区渔业资源调查与区划[M].北京:海洋出版社,1990.
[2] 金显仕,赵宪勇,孟田湘,崔毅,等.黄、渤海生物资源与栖息环境[M].北京:科学出版社,2005:144-149.
[3] 唐启升,叶懋中.山东近海渔业资源开发与保护[M].北京:中国农业出版社,1990.
[4] 程济生.黄渤海近岸海域生态环境与生物群落[M].青岛:中国海洋大学出版社,2004.
[5] 左涛,王荣,王克,高尚武.夏季南黄海浮游动物的垂直分布与昼夜垂直移动[J].生态学报,2004,24(3):524-530.
[6] 陈大刚.黄渤海渔业生态学[M].北京:海洋出版社,1991.
[7] 胡好国,万振文,袁业立.南黄海浮游植物季节性变化的数值模拟与影响因子分析[J].海洋学报,2004,26(6):74-88.
[8] 于非,张志欣,刁新源,郭景松,汤毓祥.黄海冷水团演变过程及其与临近水团的关系的分析[J].海洋学报,2006,28(5):26-34.
[9] Ludwig J A,and Reynold J F.Statistical Ecology[M].New York:John Wiely&Sons,1988.
[10] 任一平,徐宾铎,叶振江,刘元刚.青岛近海春、秋季渔业资源群落结构特征的初步研究[J].中国海洋大学学报:自然科学版,2005,35(5):792-798.
[11] Wilhm J L.Use of biomass units in Shannon’s formula[J].Ecology,1968,49:153-156.
[12] Clarke K R.Comparisons of dominance curves[J].Journal of Experimental Marine Biology and Ecology,1990,138:143-157.
[13] Clarke K R,Ainsworth M.A method of linking multivariate community structure to environmental variables[J].Mar Ecol Prog Ser,1993,92:205-219.
[14] Clarke K R,Warkwick R M.Changes in Marine Communities:An App roach to Statistical Analysis and Interp retation(2nd edition)[M].Plymouth:PRIMER-E Ltd,2001.
[15] 金显仕.山东半岛南部水域春季游泳动物群落结构的变化[J].水产学报,2003,27(1):19-24.
[16] 朱鑫华,吴鹤洲,徐凤山,等.黄渤海沿岸海域游泳动物群落多样性及其相关因素的研究[J].海洋学报,1994,16(3):102-112.
[17] 朱鑫华,吴鹤洲,徐凤山,等.黄渤海沿岸海域游泳动物群落结构时空格局异质性研究[J].动物学报,1994,40(3):241-252.
[18] 曾晓起,朴成华,姜伟,等.胶州湾及其邻近海域渔业生物多样性的调查研究[J].中国海洋大学学报:自然科学版,2004,34(6):977-982.
