长牡蛎不同地理群体选育系数量性状的比较*
2011-01-10王庆志刘士凯孔令锋
王庆志,李 琪,刘士凯,孔令锋
(中国海洋大学水产学院,山东青岛266003)
长牡蛎(Crassostrea gigas)又称太平洋牡蛎,具有生长速度快、产量高等优点,是世界上最重要的海水养殖贝类之一[1],也是我国海水养殖业的重要养殖对象。2007年中国养殖牡蛎产量达到351万t,占贝类总产量的35.3%[2],然而,养殖生产上长期近亲繁殖使长牡蛎种质资源出现抗逆性差、个体小型化、单位产量降低等现象。为保障牡蛎养殖业健康发展,需要进行长牡蛎的遗传改良和新品种的培育。近年来,美国、法国和澳大利亚相继开展了长牡蛎选择育种,并在提高生长率、抗病性等方面取得显著成效[3-6]。
虽然我国对一些重要经济贝类,如皱纹盘鲍(Haliotis discus hannai)[7]、海湾扇贝(A rgopecten irradians)[8]、虾夷扇贝(Patinopecten yessoensis)[9]的选育工作取得了一定进展,但长牡蛎的选择育种工作在我国尚属起步阶段。为了给长牡蛎的遗传改良工作提供更多的育种素材,作者于2006年2月从日本宫城和韩国釜山分别引进当地的长牡蛎养殖群体,并以这2个群体及我国山东威海的养殖群体作为繁殖亲本,通过群体选育、家系选育和杂交选育等手段开展了长牡蛎选择育种工作。
在牡蛎的选择育种工作中,颜色和壳型作为重要目标性状,一直受到遗传学家和育种学家的关注[3,11]。日本学者Imai和Sakai[12]发现,日本沿海长牡蛎的壳色、壳型、环境适应和繁殖条件随地理分布的变化存在明显的变异,且这些变异是可遗传的。Brake等[13]和Evans等[14]利用构建的全同胞家系进行分析,证明长牡蛎的壳色和外套膜色是具有较高遗传力的数量性状,并开展了相关的选育工作。Batista等[15]对长牡蛎和葡萄牙牡蛎闭壳肌痕的比较发现,闭壳肌痕的颜色属连续分布的数量性状,反映闭壳肌大小的闭壳肌痕与牡蛎的环境适应能力相关。消费者常通过壳型来快速判断牡蛎品质的优劣,一些国家根据壳型指数已制定出相关的市场标准[16]。为了查明长牡蛎不同地理群体数量性状间的差异,本文对中国、日本、韩国3个地理群体快速生长选育系F2代的重要数量性状进行了测量、统计和比较分析,旨在为进一步开展壳色、外套膜色、壳型等性状的相关选育工作提供理论基础。
1 材料与方法
1.1 材料
2007年6月,分别以中国威海、日本宫城和韩国釜山的养殖群体为亲本,构建了3个长牡蛎快速生长选育系F1代,2008年7月继续以F1代为亲本构建选育系F2代,采用常规方法进行受精、孵化、幼体培育、稚贝和成贝养成。
1.2 测量方法
2009年6月,对同海区的不同群体选育系F2代分别随机取样100个体进行相关数量性状测量。用游标卡尺测量个体的壳高、壳长、壳宽、闭壳肌痕长和闭壳肌痕高(精确度0.01 mm)(见图1),用电子天平测量个体的总重、软体部质量和壳质量(精确度0.1 g),采用Brake等[13]的方法用肉眼将个体的左壳颜色和外套膜颜色分为1~4级(见图2),采用Batista等[15]的方法用肉眼将个体的右闭壳肌痕颜色分为1~4级(见图2)。利用测量的形态性状,参考相关文献计算各项壳型指数:
出肉率(%)(ISF)=SOW/TW×100;
壳型指数A(ISA)=SW/SH[16];
壳型指数B(ISB)=SL/SH[16];
壳型指数C(ISC)=(SH+SW)/SL[17];
壳型指数D(%)(ISD)=SW/(SH+SL)×100[12];壳型指数E(ISE)=SL/(SH+SL+SW)[18];
其中SH、SL和SW分别代表壳高、壳长和壳宽(mm),SOW和TW分别代表软体部质量和总质量(g)。
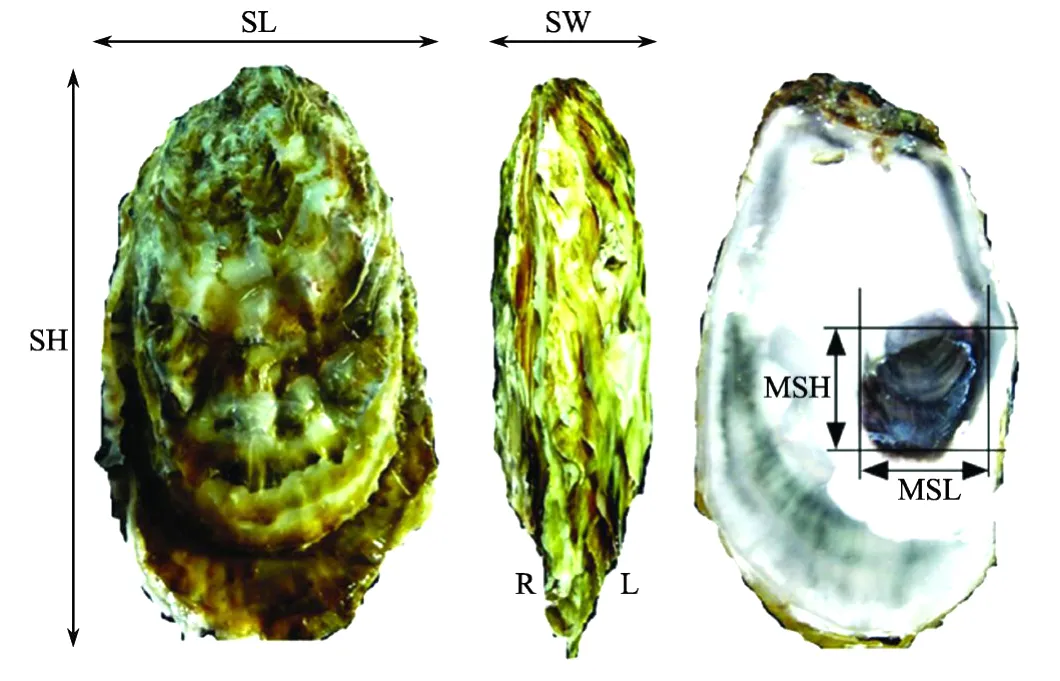
图1 长牡蛎的形态性状指标Fig.1 Shell charactersmeasured of Crassostrea gigas
1.3 数据处理与分析
为增加数据的正态性和方差均一性,在进行方差分析前,将壳长、壳高、壳宽等形态性状数据转化为常用对数。此外,为消除个体大小对形态形状的影响,通过壳重对其它形态性状的协方差分析(ANCOVA)将数据修正[15,19],再进行不同群体间的比较分析。用单因素方差分析(ANOVA)分析不同群体间各项形态指标的差异,以SN K法进行群体间的多重比较。使用R×C无序列联表的卡方检验分析不同群体间各级壳色、外套膜色及闭壳肌痕色构成比例的差异。所有统计分析均由SPSS 16.0分析软件完成,显著性水平为P<0.05。
2 结果
2.1 不同群体生长性状和壳型指数的比较
生长性状方面,日本群体与中国和韩国的群体均存在显著差异(P<0.05),而中国和韩国群体除总重差异显著外(P<0.05),其它各项指标差异均不显著(P>0.05),日本群体的所有生长性状指标均显著高于中、韩群体(见表1)。

图2 长牡蛎左壳、外套膜和闭壳肌痕的1-4级着色Fig.2 Characters pigmentation measured of C.gigas
壳型指数方面,日、韩群体的出肉率差异不显著(P>0.05),但均显著高于中国群体(P<0.05);其余指数中,除日本群体的壳型指数C显著高于中、韩群体外(P<0.05),中、韩群体的壳型指数A、B、D、E均显著高于日本群体(P<0.05)(见表2),中、韩群体的各项指数差异均不显著(P>0.05)。
2.2 个体大小校正后长牡蛎不同群体生长性状和壳型指数的比较
生长性状方面,与大小未经校正的结果相比较,校正后数据不同群体间的差异发生一定变化,日本群体的壳高、总质量和闭壳肌痕高仍显著大于中、韩群体(P<0.05),日本群体的软体部重显著大于中国群体(P<0.05),中、韩群体的壳宽显著大于日本群体(P<0.05),韩国群体的壳长显著大于日本群体(P<0.05),中、日、韩群体的闭壳肌痕长差异不显著(P>0.05),中、韩群体的各项差异均不显著(P>0.05)。
壳型指数方面,与大小数据未经校正的结果相比较,除中、日、韩群体的出肉率差异均不显著外(P<0.05),不同群体间壳型指数的差异与未校正数据的分析结果相同。

表1 长牡蛎不同群体快速生长选育系F2代的生长性状比较Table 1 Comparisons of grow th traits among different selected lines of C.gigas
表2 长牡蛎不同群体快速生长选育系F2代的壳型指数比较Table 2 Comparisons of shell shape index among different seleed lines of C.gigas

表2 长牡蛎不同群体快速生长选育系F2代的壳型指数比较Table 2 Comparisons of shell shape index among different seleed lines of C.gigas
注:同列间标注不同字母的两项间差异显著(P<0.05)。Note:The two item s with different letters in the same column are significantly different(P<0.05).
群体Stock ISF/%ISA ISB ISC ISD/%ISE中国群体CS 9.73b(2.68)0.314a(0.067)0.680a(0.113)1.98b(0.33)18.68a(3.89)0.339a(0.036)日本群体JS 11.91a(2.74)0.255b(0.074)0.531b(0.114)2.45a(0.45)16.52b(4.16)0.295b(0.037)韩国群体KS 12.04a(2.95)0.325a(0.076)0.693a(0.102)1.95b(0.28)19.21a(4.31)0.342a(0.313)

表3 个体大小校正后长牡蛎不同选育群体生长性状的比较Table 3 Comparisons of grow th traits among different selected lines of C.gigas by using size-adjusted data

表4 个体大小校正后长牡蛎不同选育群体壳型指数的比较Table 4 Comparisons of shell shape indexes among different selected lines of C.gigas by using size-adjusted data
2.3 长牡蛎颜色性状在不同选育群体中分布频率的比较
不同群体间左壳、外套膜和闭壳肌痕的4级着色程度比例组成见图3。卡方检验结果表明不同群体的左壳、外套膜和闭壳肌痕的着色差异均极显著(P<0.01)。左壳的着色程度依次为:韩国群体>日本群体>中国群体;外套膜色的着色程度依次为:日本群体>中国群体>韩国群体;闭壳肌痕的着色程度依次为:日本群体>中国群体>韩国群体。
3 讨论

图3 长牡蛎颜色性状在不同选育群体中的分布频率Fig.3 Distribution frequency of pigmentation intensity in different selected linesof C.gigas
通过比较不同群体间、不同品系间及亲本与子代间的性状差异,是育种工作者制定育种规划、选择育种亲本和判断育种效果的基本依据。本研究表明,不同群体间的主要数量性状差异显著:生长方面,日本群体的壳高和总重显著大于中、韩群体,表明日本群体具有明显的生长优势;而壳型指数方面,中国和韩国群体间无显著差异,日本群体壳型指数C显著高于中、韩群体,其余壳型指数均低于中、韩群体。由于牡蛎个体间的形态差异较大,单一性状的比较通常不能完全判断牡蛎个体间壳型的差异,而性状间的比值即壳型指数则可较好的反映这一差异,不同学者采用的壳型指数各有特点,如本文中的壳型指数D[12](又称为壳深指数)和壳型指数E[18](又称为壳凸指数)可较好的反映牡蛎壳型的凹凸情况,而壳型指数A和B[16]则可用于简单快速地区分牡蛎壳型的优劣,一些国家已利用壳型指数制定牡蛎品质的市场标准。利用个体大小校正后的数据可消除规格大小给性状间比较带来的误差[15],本研究利用协方差分析对原始数据进行校正,校正后的分析结果与原数据的分析结果基本相同,进一步证明不同群体间的数量性状存在显著差异。Imai和Sakai[12]对日本沿海不同野生长牡蛎群体的研究表明,从北海道至广岛,随着地理分布的逐渐南移,长牡蛎的个体逐渐变小,贝壳着色逐渐加深,壳型凸度逐渐增大,并且这些性状在连续数代保持稳定遗传。本研究中的3个不同地理群体,由北向南依次为日本宫城、中国威海和韩国釜山,所反映的群体间数量性状的差异也进一步验证了Imai和Sakai的结论。
在动植物的遗传育种工作中,颜色性状一直受到广泛的关注,海洋贝类的颜色丰富多彩,这些色彩中有单基因控制的质量性状,也有多基因控制连续分布的数量性状。本研究通过将长牡蛎的左壳、闭壳肌痕和外套膜的着色各分成4个等级,再进行群体间不同等级颜色构成比例的比较分析,结果表明不同群体间左壳、闭壳肌痕和外套膜的着色程度均存在显著的差异。自然状态下长牡蛎的外壳经常被各种附着物所掩盖,其色彩也容易被人们所忽视,但当长牡蛎以活体或“半壳贝”销售时,消费者对牡蛎的外壳色彩和外套膜颜色会有不同的偏好。研究表明,长牡蛎的左壳色、闭壳肌痕色和外套膜色均属于多基因控制的数量性状[13-15]。Evans等[14]利用全同胞组内相关和亲子回归估算的长牡蛎的左壳色狭义遗传力和广义遗传力分别高达0.91和0.59,表明对长牡蛎的壳色性状进行定向选育是切实可行的。贝类的颜色性状不但受遗传和环境等因素影响,还与生长等表型性状相关[20-22],以颜色性状作遗传标记,再对生长等经济性状进行定向选育,培育新的贝类品种(或品系),在我国的贝类育种工作中已有成功范例[23-25]。在本研究中,不同群体间显著的颜色性状差异进一步验证了长牡蛎颜色性状的可遗传特性,同时也为我国进一步开展长牡蛎的特色选育工作提供了良好的育种素材。
群体遗传学研究表明,贝类不同地理群体间都存在明显的基因频率和基因型差异,差异大小与空间距离和隔离状态有关,空间相距远或遗传距离远的群体间的种内杂交不存在配子亲和性和发育等方面的障碍,可直接应用推广,也可为选择育种提供基础材料[26]。利用不同遗传背景的群体间杂交可获得杂种优势,这已在皱纹盘鲍[27]和栉孔扇贝[28]等种类中得到验证。本研究中不同群体间显著的性状差异为开展长牡蛎的群体间杂交育种工作提供了参考资料。
[1] Food and Agriculture Organization(FAO).World aquaculture production of fish,crustaceans,molluscs,etc.,by principal species[EB/OL].ftp://ftp.fao.org/fi/stat/summary/default.htm.YearBooksofFisheryStatistics,2008.
[2] 农业部渔业局.中国渔业年鉴[M].北京:中国农业出版社,2007.
[3] Ward R D,English L J,Mcgoldrick D J,et al.Genetic improvement of the Pacific oyster Crassostrea gigas(Thunberg)in Australia[J].Aqua Res,2000,31:35-44.
[4] Langdon C,Evans F,Jacobson D,et al.Yieldsof cultured Pacific oysters Crassostrea gigas Thunberg improved after one generation of selection[J].Aquaculture,2003,220:227-244.
[5] Boudry P,Dégremont L,Taris N,et al.Genetic variability and selective breeding for traits of aquacultural interest in the Pacific oyster[J].Bull Aquacul Assoc Canada,2004,104:12-18.
[6] Evans S,Langdon C.Direct and indirect responses to selection on individual body weight in the Pacific oyster(Crassostrea gigas)[J].Aquaculture,2006,261:546-555.
[7] 刘晓,张国范,赵洪恩.皱纹盘鲍“中国红”品系的选育[J].动物学杂志,2003,38(4):27.
[8] Zheng H,Zhang G,Liu X,et al.Different responses to selection in two stocks of the bay scallop,A rgopecten irrad ians irradians Lamarck(1819)[J].J Exp Mar Biol Ecol,2004,313:213-223.
[9] Liang J,Zhang G,Zheng H.Divergent selection and realized heritability for grow th in the Japanese scallop,Patinopecten yessoensis Jay[J].Aqua Res,2010,41:1315-1321.
[10] 王庆志,李琪,刘士凯,等.长牡蛎幼体生长性状的遗传力及其相关性分析[J].中国水产科学,2009,16:736-743.
[11] Mahon G.Selection goals in oyster breeding[J].Aquaculture,1983,33:141-148.
[12] Imai T,Sakai S.Study of breeding of Japanese oyster,Crassostrea gigas[J].Tohoku J Agric Res,1961,12:125-171.
[13] Brake J,Evans F,Langdon C.Evidence for genetic control of pigmentation of shell andmantle edge in selected familiesof Pacific oysters,Crassostrea gigas[J].Aquaculture,2004,229:89-98.
[14] Evans S,Camara M D,Langdon C J.Heritability of shell pigmentation in the Pacific oyster,Crassostrea gigas[J].Aquaculture,2009,286:211-216.
[15] Batista FM,Ben-Hamadou R,Fonseca V G,et al.Comparative study of shell shape and muscle scar pigmentation in the closely related cupped oysters Crassostrea angulata,C.gigas and their recip rocal hybrids[J].Aqua Living Resour,2008,21:31-38.
[16] Brake J,Evans F,Langdon C.Is beauty in the eyeof the beholder?Development of a simple method to describe desirable shell shape for the Pacific oyster industry[J].J Shellfish Res,2003,22:767-772.
[17] Galtsoff P S.The American oyster Crassostrea virginica Gmelin[J].Fish Bull,1964,64:1-480.
[18] Wada K T.Genetic selection for shell traits in the Japanese pearl oyster,Pinctada fucata martensii[J].Aquaculture.1986,57(1-4):171-176.
[19] Mccoy M W,Bolker B M,Osenberg CW,et al.Size correction:comparing morphological traits among populations and environments[J].Oecologia,2006,148:547-554.
[20] Wada K T,Komaru A.Effect of selection for shell coloration on grow th rate and mortality in the Japanese pearl oyster,Pinctada fucatamartensii[J].Aquaculture,1994,125:59-65.
[21] 郑怀平,许飞,张国范,等.海湾扇贝壳色与数量性状之间的关系[J].海洋与湖沼,2008,39(4):328-333.
[22] Zheng H P,Zhang G F,Liu X.Comparison of grow th and survival of larvae among different shell color stocks of bay scallop Argopecten irradians irradians(Lamarck 1819)[J].Chinese JOceanol Limnol,2005,23(2):183-188.
[23] 闫喜武,张跃环,霍忠明,等.不同壳色菲律宾蛤仔品系间的双列杂交[J].水产学报,2008,32(6):864-875.
[24] 何毛贤.马氏珠母贝红壳品系“南科珍珠红”的培育[J].热带海洋学报,2006,25(2):58.
[25] 郑怀平,张国范,刘晓,等.不同贝壳颜色海湾扇贝(A rgopecten irradians)家系的建立及生长发育研究[J].海洋与湖沼,2003,34(6):632-639.
[26] 张国范,刘晓.关于贝类遗传改良几个问题的讨论[J].水产学报,2006,30(1):130-137.
[27] 张国范,王继红.皱纹盘鲍中国群体和日本群体的自交与杂交F1的RAPD标记[J].海洋与湖沼,2002,33(5):484-491.
[28] 刘小林,常亚青,相建海,等.栉孔扇贝中国种群与日本种群杂交一代的中期生长发育[J].水产学报,2003,27(3):193-199.
