海葵溶细胞素的生物信息学分析
2011-01-06王倩倩蔡滨欣张黎明
陆 佳,肖 良,王倩倩,蔡滨欣,张黎明
(第二军医大学 海医系 防化医学教研室,上海 200433)
海葵溶细胞素的生物信息学分析
陆 佳,肖 良,王倩倩,蔡滨欣,张黎明
(第二军医大学 海医系 防化医学教研室,上海 200433)
利用生物信息学的软件与方法对已报道的17种海葵溶细胞素的氨基酸组成、理化性质、结构特征以及系统演化关系等进行预测和分析。结果表明不同海葵溶细胞素在氨基酸组成和理化性质上具有一定相似性,但其在结构上有一定差别。以MP法和NJ法对不同海葵溶细胞素构建的系统发生树基本一致,在亲缘关系较近的7种海葵溶细胞素中可以找到6个保守序列。
海葵;溶细胞素;生物信息学
海葵(Sea anemone)属于腔肠动物门(Coelenterata)珊瑚纲(Anthozoa),是一类原始的多细胞动物。海葵毒素是一类具有溶血、细胞毒、心脏毒、膜去极化和钾离子通道阻滞等生物毒性的多肽类毒素。溶细胞素是海葵毒素中研究较为深入的一类活性组分[1-5]。本文拟利用生物信息学的软件与方法对目前已报道的17种来源不同的海葵溶细胞素的氨基酸组成和序列、理化性质、结构特征以及系统演化关系等进行预测和分析,以期为深入开展该类蛋白质的相关研究提供理论指导。
1 材料和方法
通过检索,从NCBI蛋白质数据库中筛选得到Actinoporin Or-A等17种海葵溶细胞素作为分析对象(见表1)。

表1 17种海葵溶细胞素序列
利用BioEdit等多种生物信息学软件(见表2)对17种海葵溶细胞素蛋白序列的氨基酸组成、理化性质、结构特性及分子系统进化关系等进行分析和预测。
2 结果
2.1 氨基酸组成、理化特性以及信号肽与跨膜结构域的预测
分析发现不同海葵溶细胞素的氨基酸组成具有一定相似性(见表3):①非极性氨基酸以Gly、Ala、Leu和 Val等4种含量丰富;②极性氨基酸以Lys、Tyr和Ser等3种含量丰富。③Cys含量极低,除信号肽区域外该毒素中不含有Cys。
不同海葵溶细胞素理化性质、信号肽及跨膜结构域有如下特点:①毒素相对分子质量(Mr)范围15 000~29 800,长度165~276个氨基酸,功能域长度约170个氨基酸;②除Src I和RTX-SⅡ为酸性蛋白外,其余毒素等电点均大于7,为碱性蛋白;③含有长度约20个氨基酸的信号肽,部分含信号肽的毒素中含有长约10~20个氨基酸的“Propeptide”序列;④除 Avt-1、EqTV、Src I、PsTX-20A 和 HMgⅢ这5 种海葵溶细胞素含有跨膜结构外,其余12种溶细胞素不含跨膜结构。

表2 主要分析软件[6]

表3 海葵溶细胞素的理化性质、信号肽及跨膜结构域的分析
2.2 不同海葵溶细胞素疏水性/亲水性预测
海葵溶细胞素的疏/亲水性有明显差异,可分为3组:第1组包括Actinoporin Or-A等13种溶细胞素,其结构中均有1个长约30个氨基酸的疏水区;第2组包括Avt-1、PsTX-20A和Bandaporin等3种,含有2个长度均为25个氨基酸的疏水区;第3组仅Echt 2这1种,含有2个长度分别为50和25个氨基酸的疏水区域。
2.3 二级结构预测
预测发现海葵溶细胞素二级结构相似性较高。如表4所示,除Echt2外,含量最多的是无规则卷曲,其次为β-折叠和α-螺旋,而Echt2中含量最高的是α-螺旋,其次为无规则卷曲和β-折叠。β-转角散布于溶细胞素整个蛋白分子内。
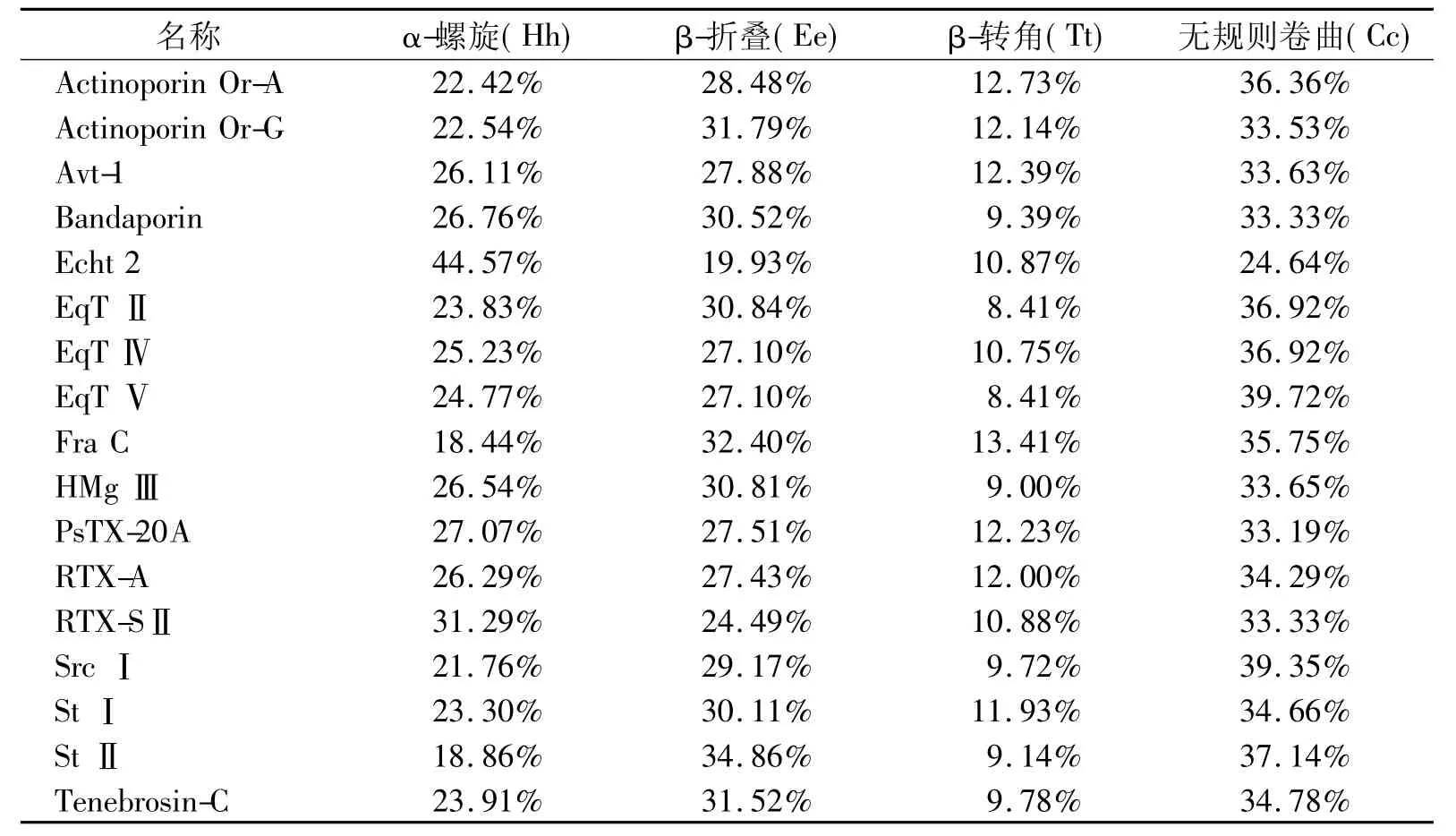
表4 不同海葵溶细胞素二级结构预测
2.4 不同海葵溶细胞素多重序列比对、分子进化及保守序列的分析
对海葵溶细胞素进行亲缘分析,发现Actinoporin Or-A、Actinoporin Or-G、Bandaporin、EqTⅡ、EqTⅣ、EqTⅤ、Fra C、HMgⅢ、RTX-A、RTX-SⅡ、StⅠ、StⅡ、Tenebrosin-C 等 13 种毒素亲缘关系较近,用MP法和NJ法分别构建了分子系统进化树(图1)。

图1 海葵溶细胞素系统进化树
结果表明,不同的建树方法,所建分子系统进化树是相似的。根据NJ法的建树结果,在Actinoporin Or-A、Actinoporin Or-G、HMgⅢ、RTX-A、RTX-SⅡ、StⅠ和 StⅡ等 7 种毒素中寻找保守序列,发现其含 VSRKIA、DNESGG、NAYFRSGTTDVILP、KALLY、YSNWW和RRADQ等6段保守序列(图2)。
2.5 不同海葵溶细胞素空间结构预测
不同海葵溶细胞素在空间结构上十分相似,多为β-折叠及α-螺旋两者间以 β-转角连接。其信号肽和“Propeptide”的序列多以α-螺旋形式存在,在毒素N端含有双亲螺旋,推测与结合膜蛋白发挥溶细胞毒性有关。

图2 海葵溶细胞素蛋白序列多重对比
3 讨论
随着传统生物学研究的不断深入和发展,生物信息学作为一门新兴学科,在生物学各领域中越来越常用。虽然生物信息学分析预测的方法是建立在对大量已知数据的分析模拟基础上的,与实验结果可能存在一定的偏差,但在课题研究前期,生物信息学的方法对揭示生物分子自身所带的信息、规律及生物大分子间的关系有重要作用[7]。本文主要利用生物信息学方法对已报道的17种海葵溶细胞素的氨基酸组成、理化性质、结构特征以及系统演化关系等进行了预测分析。结果表明,不同海葵溶细胞素在结构上存在相似性,这样的相似性是其具有相似毒性作用的物质基础。根据本文分析结果并结合相关文献报道可以看出,海葵溶细胞素为一类Mr约20 000的分泌蛋白,除个别为酸性蛋白外,多数为碱性蛋白;是一类真核生物膜孔道形成毒素(pore-forming toxin),能够与血细胞、肿瘤细胞、血小板、纤维原细胞及人工脂膜相结合,在膜上形成直径为1.0~2.0 nm的离子选择性通道[8]。在膜孔形成的过程中,毒素的正电荷与磷脂双分子层表面负电荷相互吸引靠近;毒素结构中富含Lys、Tyr、Ser等极性氨基酸的区域与细胞膜结合,N端氨基酸形成的双亲螺旋区插入膜内,达一定浓度后毒素蛋白即可结合形成三或四聚体形成阳离子通道从而发挥毒性作用[9]。若切除毒素N端的氨基酸可发现随着切除的氨基酸数目的增加,毒素蛋白的溶解性和溶血能力均降低,但其对鞘磷脂和血红细胞的结合能力却增强,说明海葵溶细胞素保守的N端与该毒素蛋白的溶解性、膜孔形成有关,但并不是膜结合所必须的[10]。综合上述分析,可以看出文献已报道的研究结果和本文利用生物信息学方法分析的结果是吻合的,提示我们可以反过来运用生物信息学方法分析、预测的结果来指导具体的实验研究工作。在对海葵溶细胞素的分析中我们发现一些保守区域,这些保守区除短肽RGD已有相关研究报道外,其余均未见报道,这些信息对我们进一步深入研究海葵溶细胞素的作用机制及相关防护措施和救治药物具有一定指导意义。可以预测,随着生物信息学理论和技术的飞速发展,利用生物信息学方法对生物大分子的结构和功能进行分析将会成为生物分子前期研究中常规的技术手段。
[1]Bellomio A,Morante K,Barlic A,et al.Purification,cloning and characterization of fragaceatoxin C,a novelactinoporin from the sea anemone Actinia fragacea[J].Toxicon,2009,54(6):869-880.
[2]Uechi G,Toma H,Arakawa T,et al.Molecular cloning and functional expression of hemolysin from the sea anemone Actineria villosa[J].Protein Expr Purif,2005,40(2):379-384.
[3]Monastyrnaya M M,Zykova T A,Apalikova O V,et al.Biologically active polypeptides from the tropical sea anemone Radianthus macrodactylus[J].Toxicon,2002,40(8):1197-1217.
[4]Wang Y,Chua K L,Khoo H E.A new cytolysin from the sea anemone,Heteractis magnifica:isolation,cDNA cloning and functional expression[J].Biochim Biophys Acta,2000,1478(1):9-18.
[5]刘 伟,王 娟,姜孝玉,等.海葵溶细胞素的研究进展[J].海洋科学,2005,29(4):56-62.
[6]张成岗,贺福初.生物信息学方法与实践[M].北京:科技出版社,2002:110-143.
[7]孙 啸,陆祖宏,谢建明.生物信息学基础[M].北京:清华大学出版社,2006:138-162.
[8]Valcarcel CA,Dalla Serra M,Potrich C,et al.Effects of lipid composition on membrane permeabilization by sticholysinⅠandⅡ,two cytolysins of the sea anemone Stichodactyla helianthus[J].Biophys J,2001,80(6):2761-2774.
[9] Hong Q,Gutierrez-Aguirre I.Two-step membrane binding by equinatioxinⅡ,a pore-forming toxin from the sea anemone,involves an exposed aromatic cluster and a flexible helix[J].J Biol Chem,2002,277(44):41916-41924.
[10] Anderluh G,Pungercar J,Krizaj I,et al.N-terminal truncation mutagenesis of equinatoxinⅡ,a pore-forming protein from the sea anemone Actinia equina[J].Protein Eng,1997,10(7):751-755.
Bioinformatic analysis on sea anemone hematoxins
LU Jia,XIAO Liang,WANG Qian-qian,CAI Bin-xin,ZHANG Li-ming
(Department of Chemical Defense Medicine,Faculty of Naval Medicine,No.2 Military Medical University,Shanghai 200433,China)
R282.76
A
1005-1678(2011)04-0332-04
2010-04-30
上海市自然科学基金(10ZR1437900)
陆 佳,女,军事预防医学专业硕士研究生;张黎明,通信作者,E-mail:lmzhang1969@yahoo.com.cn。
