苜蓿属和胡卢巴属植物的形态特征及数量分类研究
2010-12-31邓超宏崔大方羊海军李飞飞
邓超宏,崔大方,羊海军,李飞飞,方 颖
(华南农业大学 a.生命科学学院;b.林学院;c.公共基础课实验教学中心,广东 广州 510642)
苜蓿属(MedicagoL.)及其近缘属植物胡卢巴属(TrigonellaL.)等都隶属于豆科(Legum inosae)车轴草族(Tribe.Trifo lieae),在中国西部地区有较广泛的分布[1-3]。这两个属的植物含有丰富营养,是优良的饲用植物资源,在全世界范围内的农牧业生产体系中占有重要地位。
苜蓿属的一些种类与胡卢巴属植物在形态、生长习性及染色体数目等方面相似,形成大量的中间类群。许多研究结果表明,苜蓿属和胡卢巴属之间的亲缘关系极为相近,属间存在明显的过渡类群,使两属间的界限十分模糊[4-9]。这些中间的过渡类群常被称为“类苜蓿植物(m edicagoid)”或“类胡卢巴植物(trigonelloid)”,主要分布在亚洲中部的帕米尔、克什米尔、兴都库什和我国西部山地及其周围的高原地带(青海、新疆、西藏、云南等省区);它们的荚果呈扁长圆形、新月形或阔镰形,弯曲程度不一,既与典型的苜蓿属果实不同,又与典型的胡卢巴属果实相异;这些中间类群曾一度被分离出来,独立为扁蓿豆属(MelilotoidesHeister ex Fab r.)或黑荚豆属(TurukhaniaVass.)[4-5],而有学者把这些过渡类群归入胡卢巴属[6-8]或苜蓿属[9]。《中国苜蓿》一书记载中国苜蓿属植物有12种3变种6变型[3],《中国植物志》记载中国苜蓿属植物有13种1变种[10],造成这种差异的原因主要是中间类群的归属不同;国内也有一些文献专著将这些中间类群分别归入扁蓿豆属或黑荚豆属[11-12]。鉴于苜蓿属和胡卢巴属植物在分类上的混淆现状,韦直和崔大方等分别对国产的苜蓿属和胡卢巴属植物的分类问题进行了研究[13-15],但未对两属进行综合研究和比较。
在前人的研究基础上,作者广泛收集现有文献中涉及的苜蓿属和胡卢巴属种类,运用比较形态学、比较解剖学和数量分类方法,研究苜蓿属与胡卢巴属间的亲缘关系,以寻找两属间的分界依据及分属原则,解决中间过渡类群的分类归属问题,为国产苜蓿属与胡卢巴属的分类、资源评价和合理利用提供形态及解剖学依据。
1 材料和方法
1.1 材料
供种皮结构特征观察(11种)和花粉形态观察(9种)的苜蓿属和胡卢巴属种类采自中国的新疆、四川、宁夏、甘肃等省区以及澳大利亚的塔姆沃思、墨尔本、堪培拉和维多利亚等地,共53份样品,产地及凭证标本信息见表1;凭证标本保藏于华南农业大学植物标本馆(CANT)。查阅并测量保藏于中国科学院植物研究所植物标本馆(PE)、中国科学院新疆生态与地理研究所植物标本馆(XJBI)、新疆农业大学植物标本馆(XJA)、新疆畜牧科学院草原研究所植物标本室(XAG)、新疆师范大学生物系植物标本室(XJNU)、石河子大学植物标本室(SH I)、西北农林科技大学植物标本馆(WUK)及华南农业大学植物标本馆(CANT)的苜蓿属与胡卢巴属种类的标本,共计16种217份标本,测量数据用于聚类分析。
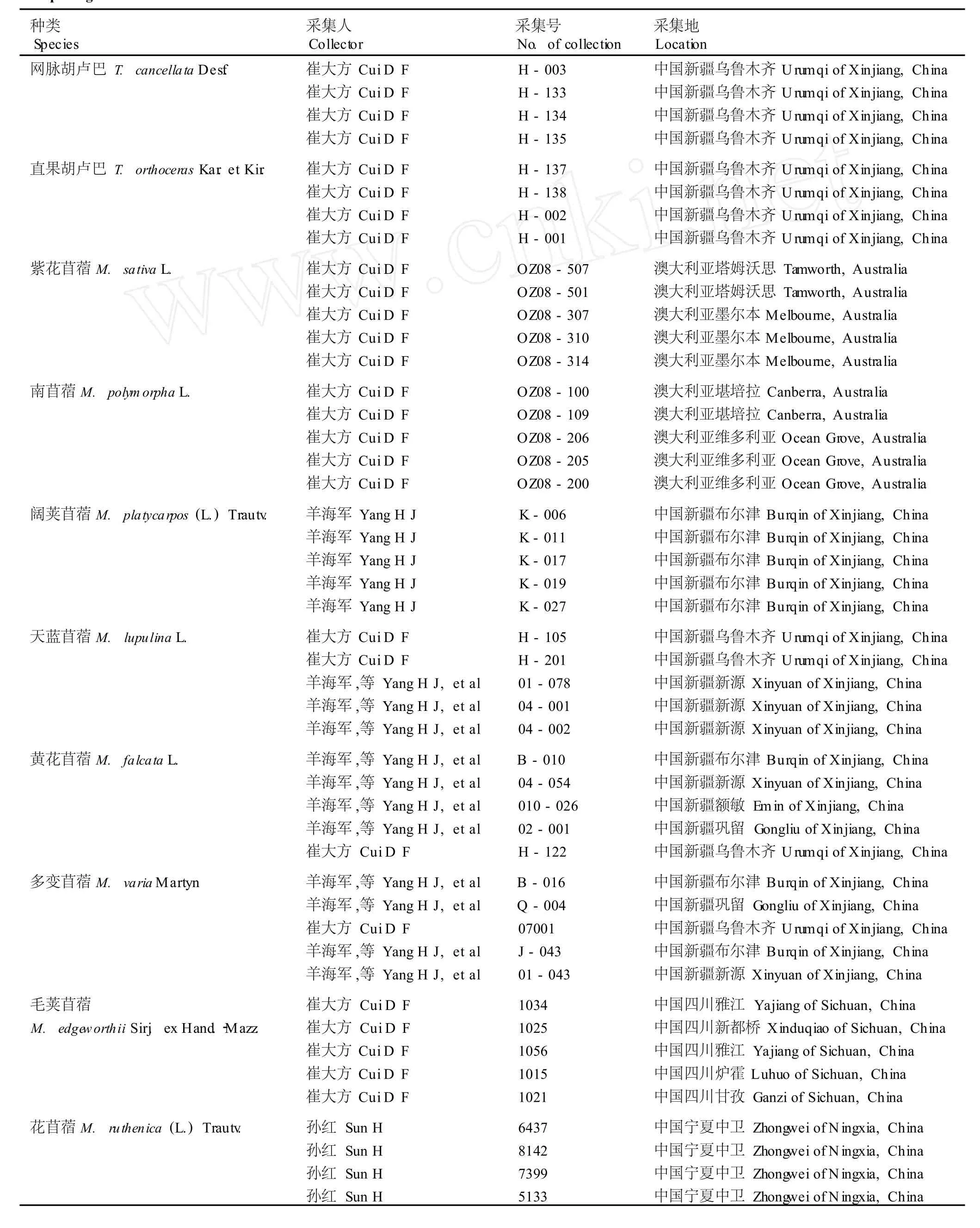
表1 用于种皮结构和花粉形态特征观察的苜蓿属和胡卢巴属种类的凭证标本基本信息Table1 Ba sic in form a tion of voucher spec im en s of Med icago L.and T rigonella L.species used for observ ing testa structure and pollenm orpholog ica l characters
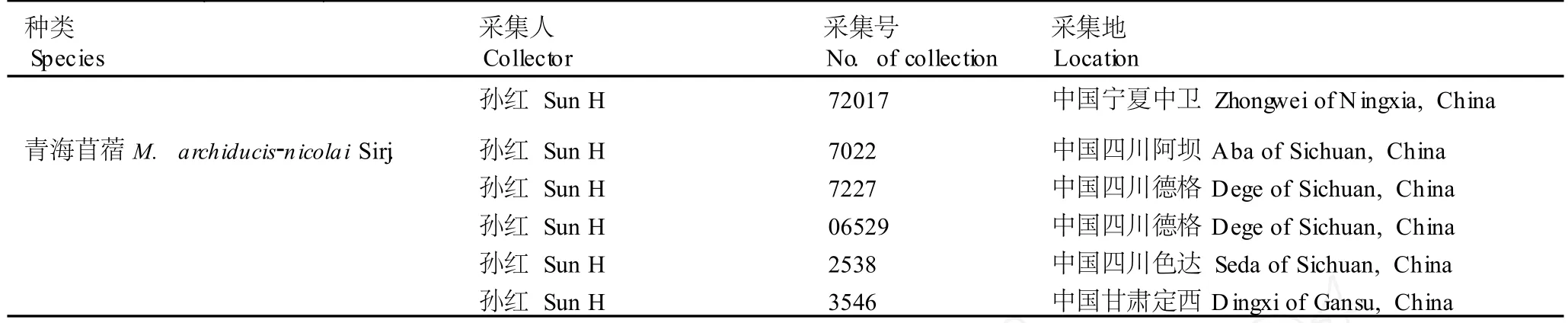
续表1 Table1(Con tinued)
1.2 方法
1.2.1 种皮结构特征观察方法 将成熟种子煮沸后剥离种皮;选取种皮中部,切成1mm×1mm的小块,用 FAA溶液固定;经体积分数50%至100%的系列乙醇梯度脱水后,用二甲苯进行透明处理,并用石蜡包埋;采用常规方法制片,切片厚度8~10μm,番红-固绿对染,中性树胶封片。用 ZeissAxioskop40型光学显微镜(德国卡尔·蔡司公司生产)以及 Motic Im agesA dvanced3.2图像处理软件进行种皮横切面结构的显微观察、拍照和测量。
1.2.2 花粉形态特征的观察方法 从标本上取下花粉,采用醋酸酐方法制片,在Motic BA200型光学显微镜(麦克奥迪实业集团有限公司生产)下进行观察和测量,每个种类观测20粒以上的花粉,测量极轴和赤道轴的长度,结果取平均值。将花粉粒均匀撒在双面胶上,经喷金镀膜后,在 Philips XL30 ESEM型环境扫描电镜(荷兰 Philips-FEI公司生产)下观察和拍照。1.2.3 聚类分析和主成分分析方法 根据《中国植物志》[10]中对苜蓿属特征描述所用的40个性状(包括宏观外部形态、果皮和种皮组织结构以及花粉微形态等)对形态性状进行测定。这些性状及编码分别为:茎形状(圆形0、四棱形1);茎被毛情况(无毛0、稀毛1、疏毛2、密毛3);托叶长;托叶形状(披针形0、戟形1、三角形2、卵形3);托叶基部形状(圆形0、耳形1、戟形2、锯齿形3);托叶边缘形状(全缘0、锯齿1);托叶被毛情况(无毛0、稀毛1、疏毛2、密毛3);托叶脉纹(不清晰0、清晰1);叶柄长;叶轴长;小叶被毛情况(无毛0、稀毛1、疏毛2、密毛3);小叶形状(倒披针形0、长椭圆形1、阔椭圆形2、倒卵形3);小叶尖形状(微凹0、截平1、钝形2);小叶边缘情况(全缘0、顶部锯齿1、1/3以上锯齿2、1/2以上锯齿3、全锯齿4);小叶基部形状(楔形0、阔楔形1);小叶长;小叶宽;小叶长宽比;小叶上表皮被毛情况(无毛0、稀毛1、疏毛2、密毛3);小叶下表皮被毛情况(无毛0、稀毛1、疏毛2、密毛3);花梗长;花梗被毛情况(无毛0、稀毛1、疏毛2、密毛3);花长;苞片长;花萼长;花萼被毛情况(无毛0、稀毛1、疏毛2、密毛3);萼齿长;萼齿形状(线形0、披针形1、锥形2);花序梗长;花序长;果实形状(肾形0、长圆形1、镰状2、螺旋状3、螺旋状带刺4);果实长;果实宽;果实长宽比;果皮被毛情况(无毛0、稀毛1、疏毛2、密毛3);花粉极轴长;花粉赤道轴长;花粉极轴与赤道轴长度比;种皮帽状结构厚度和种皮柱状结构厚度。从217份标本上获取茎、叶、花、果实和种子等器官的40个表征形态性状(包括21个数量性状和19个质量性状)的数据,每个类群分别测量15份标本,少数类群因馆藏量太少则以实测标本数量为准。
性状编码策略参照徐克学[16]的方法进行。对各性状进行编码,并以苜蓿属与胡卢巴属的16个种为运算单位,建立线性数据矩阵,然后利用 NTSYSpc version2.le软件包进行聚类分析(UPGMA)和主成分分析(PCA),分别构建树形图和二维散点图,其中聚类分析以欧氏距离为计算依据,主成分分析以相关系数为计算依据。
2 结果和分析
2.1 种皮结构特征分析
苜蓿属(9种)与胡卢巴属(2种)植物种皮的结构见图1。由图1可以看出,参试的苜蓿属与胡卢巴属种类的种皮结构基本相同,均由3层细胞构成:第1层为栅栏组织细胞,第2层为厚壁细胞,第3层为薄壁细胞。栅栏组织细胞除内切向壁不加厚外,其他方向的细胞壁均加厚,且径向壁自内向外逐渐加厚;细胞基部腔隙大,细胞核位于其中;径向壁上有不均匀加厚,自外切向壁向外突出加厚形成帽状突起。厚壁细胞均为1层,呈方形,在细胞中有1个较大的液泡腔,细胞的切向壁不加厚,但径向壁加厚。薄壁细胞位于种皮最内层,细胞层数各异,同一种植物也会因所取样品位于种皮的不同部位而出现薄壁细胞层数不同的现象。
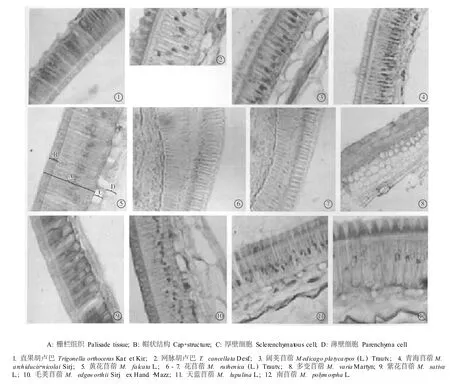
图1 苜蓿属和胡卢巴属种类种皮的横切面结构Fig.1 Tran sverse section structure of testa of Med icago L.and T rigonella L.species
供试的苜蓿属和胡卢巴属各种类的种皮和种脐形态结构也基本相同,表现为属内性状的一致性。然而,虽然供试的11个种类的种皮都是由栅栏组织细胞、厚壁细胞和薄壁细胞组成,但是不同种类种皮结构的解剖学特征存在一定的差异,具体表现在帽状结构及栅栏组织的厚度、厚壁细胞内有无沉淀物以及薄壁细胞层数等方面,各指标的实际测量结果见表2。
由表2可见,在苜蓿属和胡卢巴属的属间和属内种皮结构都有一定的差异。胡卢巴属2个种类种皮的帽状结构厚度(4.63和5.68μm)明显小于苜蓿属(7.08~8.08μm),栅栏组织厚度(20.02和24.65 μm)也明显小于苜蓿属种类(24.70~35.38μm)。属内种间种皮解剖学特征差异表现在:1)直果胡卢巴(T.orthocerasKar.et Kir.)和网脉胡卢巴(T. cancellataDesf.)种皮帽状结构厚度与栅栏组织厚度的比值和薄壁细胞层数相同(分别为0.23和1),但直果胡卢巴种皮的厚壁细胞内无沉淀物,而网脉胡卢巴种皮的厚壁细胞内有沉淀物;2)苜蓿属9个种的种皮帽状结构厚度与栅栏组织厚度比值为0.22~0.31,除阔荚苜蓿〔M.platycarpos(L.)Trautv.〕外其余种类种皮的厚壁细胞内均没有沉淀物,薄壁细胞层数为1或2层。
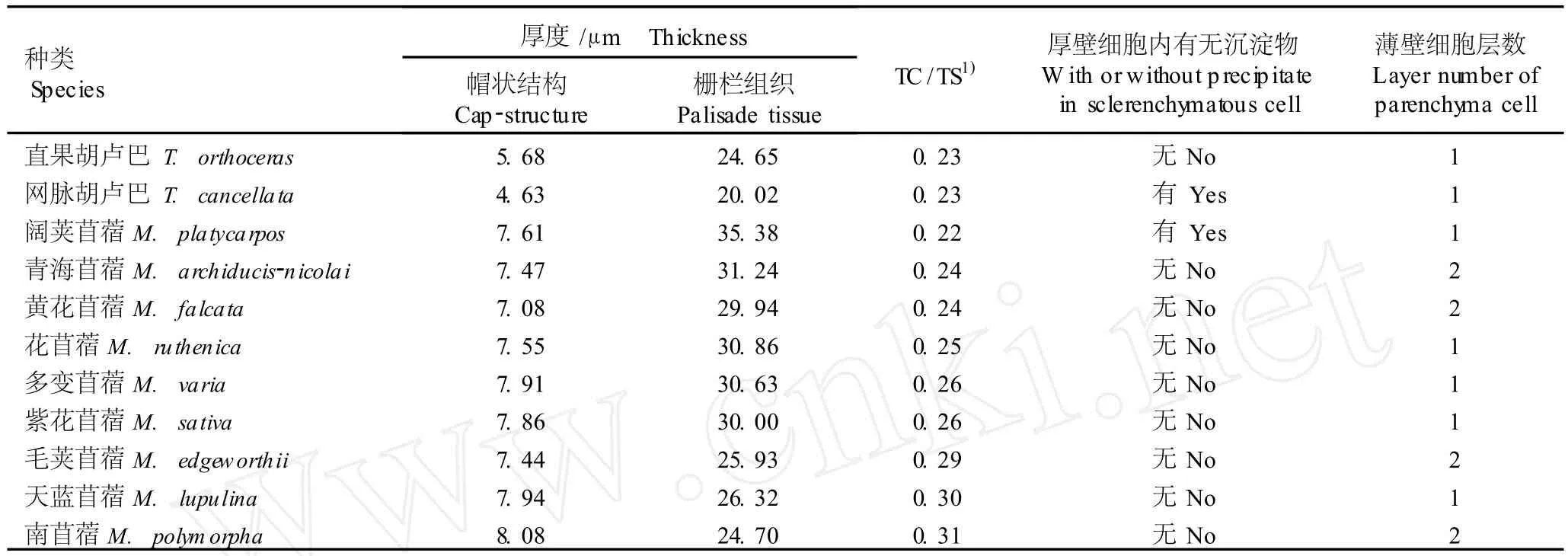
表2 苜蓿属和胡卢巴属种类种皮解剖特征的比较Table2 Com par ison of ana tom ica l character istics of testa of Med icago L.and T rigonella L.spec ies
2.2 花粉的形态特征分析
在光学显微镜下苜蓿属(7种)与胡卢巴属(2种)种类的花粉形态特征见图2,扫描电镜下花粉的形态特征及表面纹饰见图3,花粉形态的测量数据见表3。苜蓿属与胡卢巴属植物的花粉粒均为椭球形或近球形,赤道面观为椭圆形,极面观为三裂圆形;花粉粒的大小差别不大:极轴平均长22.67~34.67μm,赤道轴平均长17.34~31.60μm;花粉粒均具三孔沟,孔处有或无外突;花粉粒表面纹饰较简单,光学显微镜下多呈平滑或近平滑,扫描电镜下表面多呈小穴状。
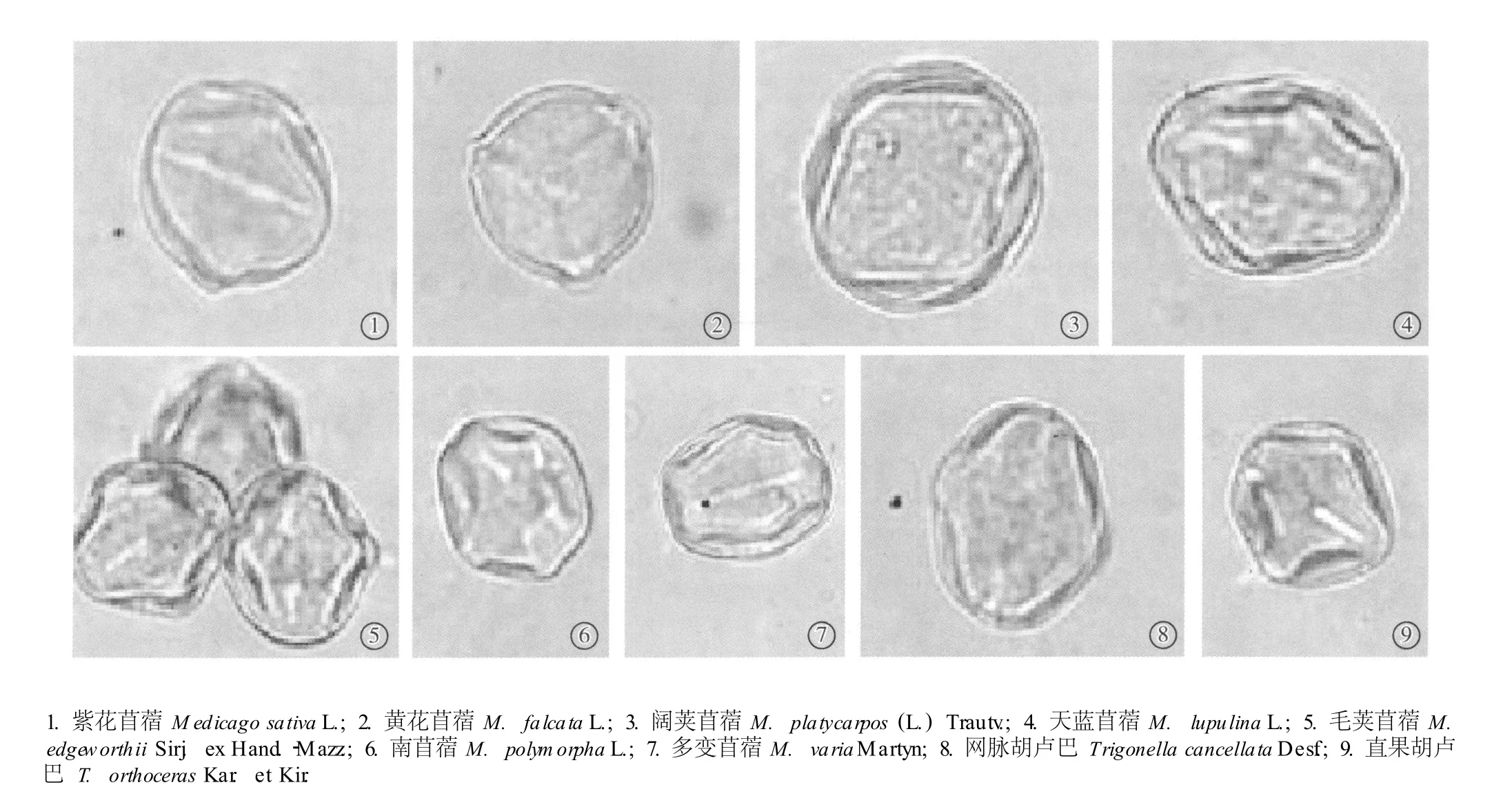
图2 光学显微镜下苜蓿属和胡卢巴属种类的花粉粒形态Fig.2 Pollenm orphology of Med icago L.and T rigonella L.spec ies under ligh tm icroscope
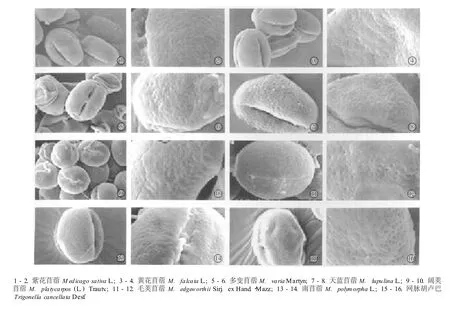
图3 扫描电镜下苜蓿属与胡卢巴属种类的花粉粒形态Fig.3 Po llen m orpho logy of Med icago L.and Trigonella L.spec ies under scann ing electron m icroscope
表3 苜蓿属和胡卢巴属种类花粉性状的测定结果(±SD)Table3 Determ ina tion resu lts of po llen character of Med icago L.and T rigonella L.spec ies(±SD)
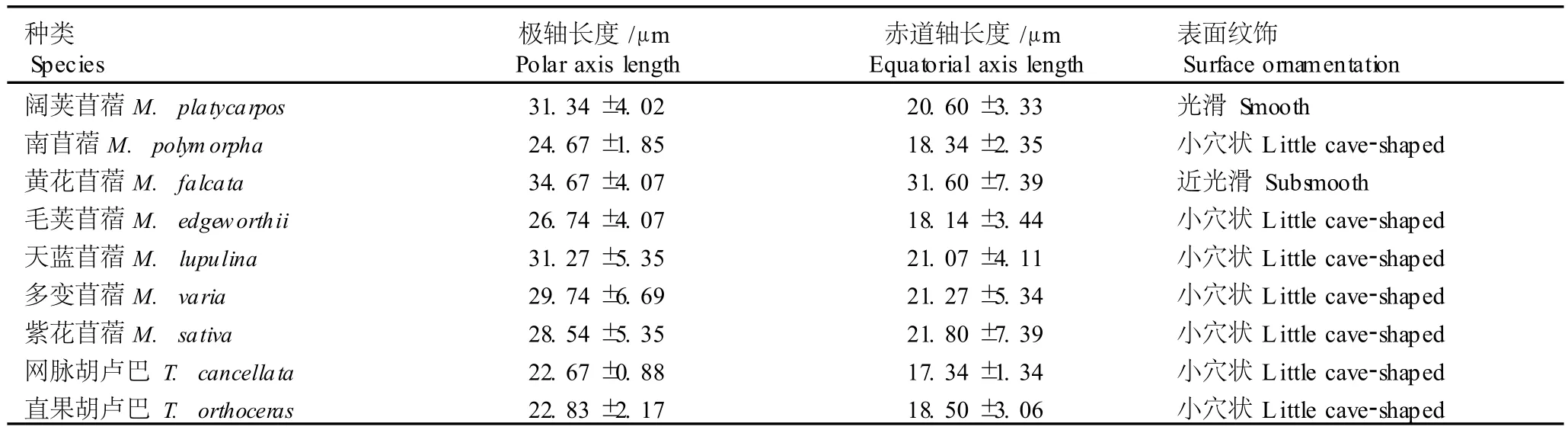
表3 苜蓿属和胡卢巴属种类花粉性状的测定结果(±SD)Table3 Determ ina tion resu lts of po llen character of Med icago L.and T rigonella L.spec ies(±SD)
种类Species极轴长度/μm Polar axis length赤道轴长度/μm Equatorial axis length表面纹饰Surface ornamentation阔荚苜蓿M.pla tycarpos 31.34±4.02 20.60±3.33 光滑Smooth南苜蓿M.polym orpha 24.67±1.85 18.34±2.35 小穴状L ittle cave-shaped黄花苜蓿M.fa lca ta 34.67±4.07 31.60±7.39 近光滑Subsmooth毛荚苜蓿M.edgew orthii 26.74±4.07 18.14±3.44 小穴状L ittle cave-shaped天蓝苜蓿M.lupu lina 31.27±5.35 21.07±4.11 小穴状L ittle cave-shaped多变苜蓿M.varia 29.74±6.69 21.27±5.34 小穴状L ittle cave-shaped紫花苜蓿M.sa tiva 28.54±5.35 21.80±7.39 小穴状L ittle cave-shaped网脉胡卢巴T.cancella ta 22.67±0.88 17.34±1.34 小穴状L ittle cave-shaped直果胡卢巴T.orthoceras 22.83±2.17 18.50±3.06 小穴状L ittle cave-shaped
2.3 基于形态性状的聚类分析结果
以苜蓿属与胡卢巴属的16个种为运算单位、基于40个形态性状(包括宏观外部形态、果皮和种皮组织结构以及花粉微形态等),利用 UPGMA法进行聚类分析,结果见图4。在欧氏距离10.85处,16个种类被划分为2组:A组和B组,具有属界特征。其中, A组包含了直果胡卢巴、单花胡卢巴(T.monanthaC. A.Meyer)、胡卢巴(T.foenum-graecumL.)、网脉胡卢巴和弯果胡卢巴(T.arcuataC.A.Meyer)5个种,均为传统分类中的胡卢巴属种类,最明显的特征为荚果呈线形或细长的圆锥状;B组包含了紫花苜蓿(M. sativaL.)、多变苜蓿(M.variaMartyn)、花苜蓿〔M.ruthenica(L.)Trautv.〕、毛荚苜蓿(M.edgeworthiiSirj.ex Hand.-Mazz.)、阔荚苜蓿、南苜蓿(M. polymorphaL.)、小苜蓿〔M.minima(L.)Grufb.〕、黄花苜蓿(M.falcataL.)、青海苜蓿(M.archiducisnicolaiSirj.)、天蓝苜蓿(M.lupulinaL.)和克什米尔苜蓿〔M.cachemiriana(L.)D.F.Cui〕,全部为苜蓿属或类苜蓿属种类。由此可见,形态上介于苜蓿属和胡卢巴属之间的类苜蓿属种类更适合归属于苜蓿属。
在欧氏距离9.32处,B组种类可进一步划分成3个亚组。其中,B1亚组有5种,种类最多,包含花苜蓿、毛荚苜蓿、阔荚苜蓿、青海苜蓿和克什米尔苜蓿,共同特征为荚果呈扁平、长圆形,与传统的阔荚组相一致;B2亚组包含紫花苜蓿、多变苜蓿和黄花苜蓿,特征为多年生、荚果由镰状弯曲到1~4圈螺旋卷曲;B3亚组包含天蓝苜蓿、小苜蓿和南苜蓿,特征为1~2年生、植株被毛、荚果形状为肾形或盘状具棘刺。

图4 基于形态性状的苜蓿属与胡卢巴属植物的聚类分析(UPGMA)结果Fig.4 Resu lt of cluster ana lysis(UPGMA) of Med icago L.and T rigonella L.species based onm orpholog ica l characters
2.4 基于形态性状的主成分分析结果
贡献率是表示主成分在整个数据分析中所承担的主成分意义所占的比例。取前m个主成分来代替原来全部性状所提供的信息时,累积贡献率的大小可体现出这种取代的可靠性,累积贡献率越大,可靠性越大;反之,可靠性越小。一般情况下,累积贡献率在70%以上,事物的基本面貌可以得到反映[16]。基于40个形态性状的苜蓿属和胡卢巴属种类的主成分分析结果见表4。由表4可见,前3个主成分的累积贡献率仅为57.50%,直至第5个主成分的累积贡献率才达到70%以上,说明各主成分的增值比较缓慢。
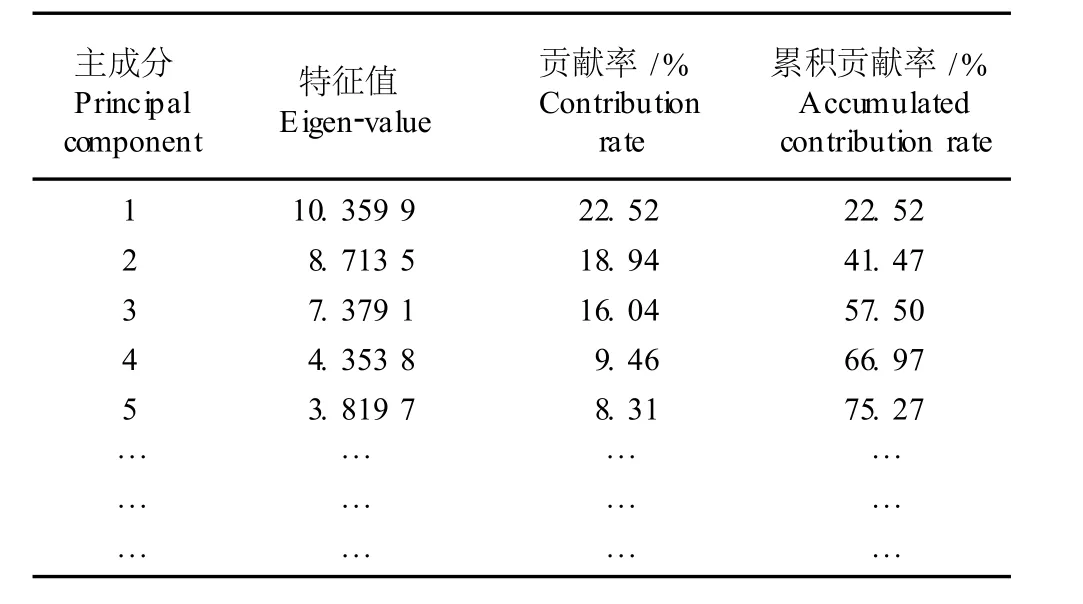
表4 苜蓿属与胡卢巴属形态性状的主成分分析结果Table4 Resu lt of p r inc ipa l com ponen t ana lysis of m orpho log ica l characters of Med icago L.and T rigonella L.
根据对各性状绝对权重值的分析可见,在第1主成分特征向量中,小叶下表皮被毛情况(0.8597)、小叶宽(-0.7992)、茎被毛情况(0.7861)、花萼被毛情况(0.7838)、托叶被毛情况(0.7280)和小叶上表皮被毛情况(0.7026)等性状的绝对权重值都在0.7以上,说明这些性状的信息负荷量最大,对分类最为重要,是区分苜蓿属与胡卢巴属植物的主要性状。在第2主成分特征向量中,果实长宽比(0.9152)、种皮帽状结构厚度(-0.8801)、花萼长(0.7789)等性状的绝对权重值也在0.7以上,说明这些性状信息负荷量也很大,对分类较重要。在第3主成分特征向量中,萼齿长(-0.8237)、小叶长宽比(-0.7806)、苞片长(-0.7655)、小叶形状(0.7379)、花序长(-0.6457)和花粉极轴与赤道轴长度的比值(0.6603)等性状的绝对权重值在0.6以上,说明这些性状对苜蓿属与胡卢巴属植物的分类也有较大的作用。
基于前2个主成分构建的苜蓿属和胡卢巴属16个种类的OTU散点图见图5。在散点图上可以很明显地看出供试的苜蓿属与胡卢巴属16个种类有4个不同的表征类群,即:胡卢巴组、天蓝苜蓿组、阔荚苜蓿组和紫花苜蓿组,并且这些类群与聚类分析划分出的分类群是一致的。说明在前2个主成分中,根据绝对权重值较高的这些性状可以进行苜蓿属与胡卢巴属植物的分类和分组。例如,通过茎、叶及花的被毛状况可以区分出1年生苜蓿类群;根据果实长宽比这一性状可以区分胡卢巴类群和阔荚类群。
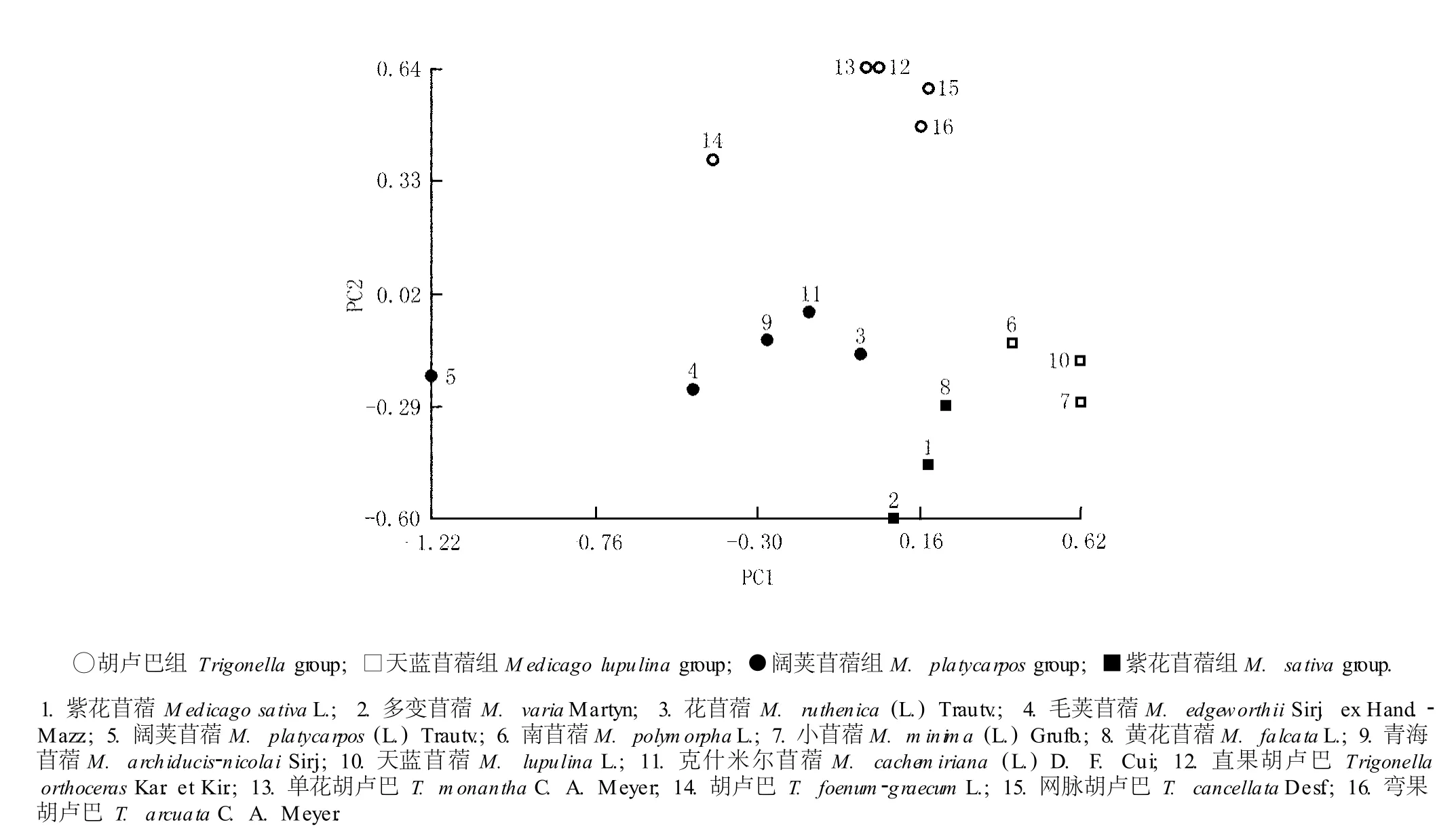
图5 基于第1和第2主成分(PC1和 PC2)构建的苜蓿属与胡卢巴属植物的二维散点图Fig.5 Two-d im en siona l sca tter d iagram of Med icago L.and T rigonella L.species based on the firstand the second p r inc ipa l com ponen ts(PC1 and PC2)
3 讨论和结论
3.1 苜蓿属和胡卢巴属植物形态特征的分类学意义
苜蓿属和胡卢巴属植物在果皮和种皮结构特征上具有一致性,这一现象在一定程度上反映了这2个属的种类间具有较近的亲缘关系,也说明单独使用果皮或种皮性状进行分类的意义不大,但是种皮帽状结构具有一定的分类价值,并成为第2主成分的性状之一。因而,若要全面分析苜蓿属和胡卢巴属植物各器官性状之间的联系,果皮和种皮性状在分类中的意义是不能被忽视的。
苜蓿属和胡卢巴属植物花粉的形态特征比较相似,外壁纹饰均较简单。马剑敏在光学显微镜和扫描电镜下观察了苜蓿属11个种类的花粉形态,认为苜蓿属属内花粉特征有较高的一致性,花粉性状分化程度小,各个种之间具有较近的亲缘关系[1];在本研究中,胡卢巴属的网脉胡卢巴和直果胡卢巴的花粉形态与供试的苜蓿属植物的花粉形态相近,区别在于花粉体积略小。由此推断,苜蓿属和胡卢巴属植物花粉性状的分类价值不大。
从40个形态性状的主成分分析结果看,前3个主成分的累积贡献率仅有57.50%,直至第5个主成分的累积贡献率才达到70%以上,表明各性状的贡献率较分散,累积贡献率增长不明显,说明国产苜蓿属和胡卢巴属种类在演化过程中性状变异具有多样性,但前2个主成分中具有较高绝对权重值的一些性状可用于苜蓿属和胡卢巴属的类群划分。
3.2 苜蓿属和胡卢巴属植物的亲缘关系分析
苜蓿属和胡卢巴属植物具有极其相似的特征,二者之间的分类界限不明显,它们有可能是由共同的祖先进化而来的,并在进化过程中演化成4个分支:一支趋向于胡卢巴属,另外三支趋向于苜蓿属的3个亚属。基于40个形态性状、采用聚类分析方法可将供试的苜蓿属和胡卢巴属16个种类划分为4组,可分别指向胡卢巴属以及苜蓿属的阔荚亚属(Subgen.PlatycarposD.F.Cui)、镰荚亚属(Subgen.MedicagoTu tin)和天蓝苜蓿亚属(Subgen.LupulinaGressh.)。其中,紫花苜蓿、多变苜蓿以及黄花苜蓿在欧氏距离5.88处聚在一起,说明它们之间的表征相似性较高;而紫花苜蓿和黄花苜蓿可以在自然条件下杂交,产生的后代为多变苜蓿,聚类分析结果也证明了它们之间具有很近的亲缘关系,应将它们归为镰荚亚属。阔荚类植物具有相似的荚果类型,其归属一直是苜蓿属与胡卢巴属植物分类的争议焦点,聚类分析结果表明,阔荚类群与苜蓿属其他种的亲缘关系较其与胡卢巴属植物的亲缘关系近,应归于苜蓿属。克什米尔苜蓿原归为胡卢巴属植物(T.cachemirianaL.),后经作者的详细研究和分析后重新归入苜蓿属中,该种具有以下特征:小叶为倒卵形,近等大,长(5)7~12 mm,宽(4)6~8 mm,先端钝圆至截平;荚果长圆形,长(10)12~15mm,宽4mm,扁平。这些特征都跟苜蓿属的阔荚类群的相应性状极为相似,在聚类分析图中,克什米尔苜蓿也被归在阔荚类群中,因此应将该种划归到苜蓿属的阔荚亚属中。胡卢巴属是苜蓿属的近缘属,在聚类图上可以看到胡卢巴属类群与苜蓿属的3个类群的关系都比较远。Sm all[7]曾提出要将胡卢巴归属到角形果组并合并到苜蓿属中,而本研究中供试的网脉胡卢巴、单花胡卢巴、直果胡卢巴和弯果胡卢巴都属于角形果组,聚类分析结果也显示它们之间有较近的亲缘关系,而与非角形果组的胡卢巴有明显区分,所以这两组植物能否归并到苜蓿属中还需进一步的深入研究。
3.3 国产苜蓿属和胡卢巴属植物的检索表
主成分分析结果显示,果实的长宽比、小叶的长宽比、花萼长、花萼被毛状况、茎和小叶被毛状况、小叶尖形状、花序长等性状对苜蓿属与胡卢巴属植物的分类具有重要意义。根据上述性状,编写出苜蓿属和胡卢巴属植物的检索表。

供试苜蓿属和胡卢巴属16个种类的检索表

6.荚果扁平、长圆形;多年生(阔荚亚属 Subgen.Pla tycarpos D.F.Cui)7.荚果长圆形至半月形,宽4mm以上8.荚果和植株均密被毛毛荚苜蓿M.edgew orthii Sirj.ex Hand.-Hazz.………………………………………………………8.荚果几无毛;植株被微毛或无毛9.荚果宽7~9mm;小叶较大;茎无毛 阔荚苜蓿M.platycarpos(L.)Trautv.………………………………………………9.荚果宽6mm以下;小叶较小;茎或多或少被柔毛10.花序松散,具花4~5朵;小叶倒卵形至圆形,托叶戟形 青海苜蓿M.arch iducis-n ico la i Sirj.………………………10.花序紧列,具花6~9朵;小叶倒披针形、楔形至线形,托叶披针形 花苜蓿M.ru thenica(L.)Trau tv.……………7.荚果长圆形,直,宽约4mm,顶端具喙 克什米尔苜蓿M.cachem iriana(L.)D.F.Cui………………………………………6.荚果镰状弯曲至螺旋状盘曲、旋转呈球形或肾形弯曲;一、二年生或多年生11.多年生(镰荚亚属 Subgen.M ed icago Tutin)12.荚果镰状弯曲达半圈左右;花冠纯黄色黄花苜蓿M.fa lca ta L.………………………………………………………12.荚果螺旋状盘曲;花冠紫色或其他杂色13.荚果镰状或螺旋状盘曲1.0~2.5圈,中央有孔;花冠紫色或其他杂色 多变苜蓿M.varia Martyn…………………13.荚果螺旋状盘曲2~4圈,中央无孔或近无孔;花冠纯紫色 紫花苜蓿M.sa tiva L.…………………………………11.一、二年生(天蓝苜蓿亚属 Subgen.Lupu lina Gressh.)14.荚果旋转呈球形,果皮具刺15.叶片无毛或近无毛;荚果直径5~10mm南苜蓿M.po lym orpha L.…………………………………………………15.叶片明显被毛;荚果直径小于4.5mm 小苜蓿M.m inim a(L.)Grufb.………………………………………………14.荚果肾形弯曲,果皮无刺天蓝苜蓿M.lupu lina L.………………………………………………………………………
[1]马剑敏.西北地区苜蓿属(MedicagoL.)的种类和分布[J].河南师范大学学报:自然科学版,1991(3):61-65.
[2]崔大方,田允温,闵继淳,等.新疆苜蓿属植物的分类研究[M]∥于兆英.西北地区现代植物分类学研究:第一卷.北京:科学技术文献出版社,1992:43-57.
[3]耿华珠.中国苜蓿[M].北京:中国农业出版社,1995:10-24.
[4]Grossheim A A.Flora of the USSR:Vo l.11[M].Moscow-Leningrad:USSR,1945:129-176.
[5]Go loskokov V P.Flora of Kazakhstana:Vo l.5[M].A lm a-A ta: Izd-vo A kadem iinauk KazakhskoǐSSR,1961:30-50.
[6]Lesins K A,Lesins I.GenusMedicago(Legum inosae),a Taxogenetic Study[M].Hague:D rWJunk Pub lisers,1979.
[7]Sm allE,Cromp ton CW,BrookesBS.The taxonom ic value of floral characters in tribe Trigonelleae(Legum inosae),with special reference toMedicago[J].Canadian Journal of Botany,1981,59(9):1578-1598.
[8]Sm all E,JompheM.A synopsis of the genusMedicago(Legum inosae)[J].Canadian Journal of Botany,1989,67(11):3260-3294.
[9]Bena G.Mo lecu lar phylogeny suppo rts the morpho logically based taxonom ic transfer of the“m edicagoid”Trigonellaspecies to the genusMedicagoL.[J].Plant System atics and Evo lution,2001,229(3/4):217-236.
[10]中国科学院中国植物志编辑委员会.中国植物志:第四十二卷第二分册[M].北京:科学出版社,1994:304-328.
[11]内蒙古植物志编辑委员会.内蒙古植物志:第二卷[M].呼和浩特:内蒙古人民出版社,1978.
[12]新疆八一农学院.新疆植物检索表:第三册[M].乌鲁木齐:新疆人民出版社,1983.
[13]韦 直.胡卢巴属与苜蓿属分类界限[C]∥中国植物学会.中国植物学会65周年年会学术报告及论文摘要汇编.北京:中国林业出版社,1998:137.
[14]崔大方,丘安经.中国苜蓿属的起源、演化与地理分布[C]∥中国植物学会.中国植物学会65周年年会学术报告及论文摘要汇编.北京:中国林业出版社,1998:58-59.
[15]崔大方,羊海军,赵业彬,等.紫花苜蓿复合体(Medicago sativacomp lex)叶片形态特征及数量分类研究[J].植物资源与环境学报,2010,19(3):1-9.
[16]徐克学.数量分类学[M].北京:科学出版社,1994.
