湖南崀山丹霞地貌区维管植物区系特征分析及对比
2010-12-31吴文文袁穗波刘帅成钟志凌旷建军罗仲春
吴文文,袁穗波,刘帅成,钟志凌,旷建军,罗仲春
丹霞地貌是中国著名地质学家陈国达在对广东省韶关市仁化县丹霞白垩系“红层”特异的地形景观进行研究后首次提出的一种特殊地貌[1]。湖南崀山丹霞盆地形成于白垩纪,丹霞地貌成型于第三纪晚期和第四纪,发育类型齐全、形态结构完整、规模宏大,在国内外丹霞地貌区中具有典型的代表性[2]。丹霞地貌由许多的孤立丹霞单体组成,正地貌悬崖峭壁,负地貌陡峭深切;丹霞山体呈孤岛状[3],这些孤岛对植物区系有深刻的影响,使其组成既具有地带性特点又出现特异现象。目前,国内对丹霞地貌植被特征的研究尚不多见,少量的研究仅见于对其植物资源利用[4]和野生藤本植物区系特征[5]的分析,缺乏对该地貌维管植物区系特征的研究。
作者对湖南崀山丹霞地貌区维管植物区系特征进行了调查研究,并与同县域的舜皇山自然保护区(非丹霞地貌)的植物区系特征进行了比较和分析,以期为认识湖南崀山丹霞地貌区植被特征与自然遗产价值提供理论基础。
1 研究区自然概况和研究方法
1.1 研究区自然概况
湖南崀山丹霞地貌区是以丹霞地貌景观为特点的风景名胜区,地理位置、气候、地貌、岩石、土壤、海拔和地带性植被等相关信息见文献[5]。
舜皇山非丹霞地貌区位于湖南省新宁县东南部,具体地理位置为北纬26°15′06″~26°55′22″、东经110°28′53″~111°18′34″;核心区面积9073.4 hm2;年均气温15.0℃;年均降水量1360.6mm。中低山地貌,岩石为花岗岩,土壤为红壤、黄壤和山地草甸土[6]。
1.2 研究方法
于2008年3月至12月采用线路与标准地相结合的方法对崀山丹霞地貌区的维管植物区系进行调查,线路调查按照溪沟或沟谷、山脊、山坡、开阔地顺序进行。在八角寨、骆驼峰、牛鼻寨、林家寨、紫霞洞、天生桥、天一巷和黄背8个景区以及穿越景区的扶夷江和湘桂交接溪谷等区域共设置12条调查线路,其中,八角寨景区和骆驼峰景区各设置2条调查线路,其他6个景区以及穿越景区的扶夷江和湘桂交接溪谷各设1条调查线路,12条调查线路总长34600m。在线路调查的基础上,选择有代表性的典型地段设立标准地进行样方调查[7-9],设植物群落标准地样方76个,每个样方面积为600m2,面积不足600m2的群落则根据优势种的分布设面积300~400m2的样方。
于2005年7月至10月,采用同样方法在舜皇山自然保护区的核心区和试验区设立14条调查线路,调查线路总长41000m,设标准地样方67个,每个样方面积为600m2。
分别记录调查时间和地点、样方内植物的种类和生长势以及土层厚度、坡度、坡向和坡位;采集植物标本并进行标本分类和鉴定[10-11];编制植物名录并进行调查资料的统计和植物区系的分析。
以上述调查数据为依据,参照罗仲春[12]的统计和分析方法,分别进行植物区系类群统计,并根据2006年吴征镒对中国种子植物分布区类型的划分原则与方法[13],对数据进行转换与统计,确定分布区类型,以作进一步的对比和分析。
分别采用秦仁昌系统和郑万钧系统对蕨类植物和裸子植物进行分类鉴定;采用哈钦松系统对崀山丹霞地貌区分布的被子植物进行分类鉴定,而舜皇山非丹霞地貌区分布的被子植物则采用恩格勒系统进行分类鉴定,故2个地貌区间科级类群不作详细对比。
2 结果和分析
2.1 崀山丹霞地貌区维管植物区系特征分析
2.1.1 维管植物区系的组成 湖南崀山丹霞地貌区维管植物区系组成的调查结果见表1。从表1中可以看出:崀山丹霞地貌区共有野生维管植物185科689属1464种(含变种),文献报道中国共有维管植物371科3347属28905种[13-14],因而,崀山丹霞地貌区维管植物科、属和种总数分别占中国维管植物科、属和种总数的49.87%、20.59%和5.06%。其中,蕨类植物有25科49属84种,分别占崀山丹霞地貌区维管植物科、属、种总数的13.51%、7.11%和5.73%;裸子植物有5科8属8种,分别占崀山丹霞地貌区维管植物科、属、种总数的2.70%、1.16%和0.55%;被子植物有155科632属1372种,分别占崀山丹霞地貌区维管植物科、属、种总数的83.78%、91.73%和93.72%。被子植物中双子叶植物有1143种(实际调查统计数),占崀山丹霞地貌区维管植物总种数的78.07%,处优势地位。
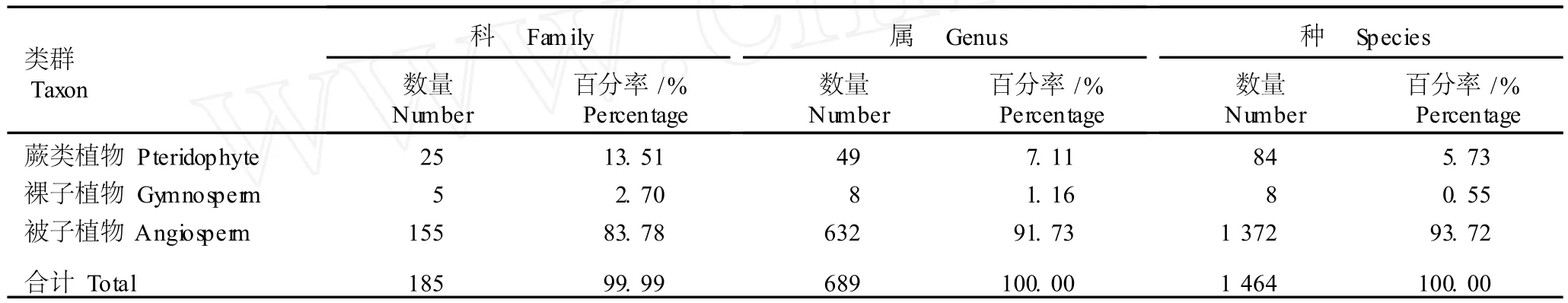
表1 湖南崀山丹霞地貌区维管植物区系组成Table1 Flora com position of vascu lar p lan ts in“Danx ia”geom orpholog ica larea of Langshan Moun ta in in Hu’nan
2.1.2 主要类群分析 崀山丹霞地貌区蕨类植物中含10种以上的科为水龙骨科(Po lypodiaceae,9属14种),含5~9种的科有卷柏科(Selaginellaceae,1属6种)、中国蕨科(Sinop teridaceae,5属6种)、凤尾蕨科(Pteridaceae,1属5种)、蹄盖蕨科(A thyriaceae,4属9种)、金星蕨科(Thelyp teridaceae,5属6种)、鳞毛蕨科(D ryop teridaceae,3属9种 )和铁角蕨科(A sp leniaceae,1属6种),其他科所含种类均在5种以下,表明湖南崀山丹霞地貌区的蕨类植物缺乏大科。
崀山丹霞地貌区分布的裸子植物均为较小科,其中,柏科(Cup ressaceae)含3属3种、红豆杉科(Taxaceae)含2属2种,松科(Pinaceae)、杉科(Taxodiaceae)和三尖杉科(Cephalotaxaceae)均仅含1属1种。
崀山丹霞地貌区被子植物中含30种以上的科有12个,仅占该地貌区被子植物总科数的7.74%。这12个科共包括233属543种,占该地貌区被子植物总属数的36.87%、总种数的39.58%。其中,含50种以上的科为蔷薇科(Rosaceae,21属60种)、菊科(Compositae,44属79种)、蝶形花科(Pap ilionaceae,25属52种)和禾本科(Gram ineae,38属50种 );含40~49种的科为兰科(O rchidaceae,20属41种)和唇形科(Labiatae,22属49种);含30~39种的科包括樟科(Lauraceae,10属36种)、山茶科(Theaceae,5属32种)、壳斗科(Fagaceae,6属38种 )、荨麻科(U rticaceae,10属33种)、茜草科(Rubiaceae,21属34种)和莎草科(Cyperaceae,11属39种)。这些科中,除菊科、荨麻科、莎草科和茜草科所含的种类为林下、林缘组成成分外,其余科所含的种类均为该区域植物群落的建群种。说明含30种以上的科在崀山丹霞地貌区植物区系中起着重要的作用。
湖南崀山丹霞地貌区共有被子植物632属,其中,含6种(变种)及以上的属有52个,共包含了427种(变种),其属数和种数分别占该区域被子植物总属数和总种数的8.23%和31.12%,这些属是湖南崀山丹霞地貌区被子植物区系中的重要组成类群。含15种以上的属只有薹草属(CarexL.),含15种。含10~14种的属有11个,其中,含10种的属有青冈属(CyclobalanopsisOerst.)、楼梯草属(ElatostemaJ.R. etG.Forst.)、山胡椒属(LinderaThunb.)和山矾属(SymplocosJacq.);含11种的属有悬钩子属(RubusL.)和柃木属(EuryaThunb.);含12种的属有冬青属(IlexL.)、槭属(AcerL.)和忍冬属(LoniceraL.);含13种的属有山茶属(CamelliaL.)和堇菜属(ViolaL.)。含5~9种的属有40个(共288种),其中,含6种的属有樟属(CinnamomumTrew)、紫堇属(CorydalisDC.)、桂樱属(LaurocerasusTourn.ex Duh.)、石楠属(PhotiniaL ind l.)、蔷 薇 属(RosaL.)、地 锦 属(ParthenocissusPlanch.)、紫金牛属(ArdisiaSw artz)、柿属(DiospyrosL.)、鹅绒藤属(CynanchumL.)、半蒴苣苔属(HemiboeaC larke)、眼子菜属(PotamogetonL.)、灯心草属(JuncusL.)和菝葜属(SmilaxL.);含7种的属有榕属(FicusL.)、苎麻属(BoehmeriaJacq.)、崖豆藤属(MillettiaWightetA rn.)、花椒属(ZanthoxylumL.)、鼠李属(RhamnusL.)、金丝桃属(HypericumL.)、胡颓子属(ElaeagnusL.)、珍珠菜属(LysimachiaL.)、蒿属(ArtemisiaL.)、鼠麴草属(GnaphaliumL.)、薯蓣属(DioscoreaL.)和虾脊兰属(CalantheR.Br.);含8种的属有毛茛属(RanunculusL.)、含笑属(MicheliaL.)、南蛇藤属(CelastrusL.)、卫矛属(EuonymusL.)、葡萄属(VitisL.)、蛇葡萄属(AmpelopsisMichx.)、猕猴桃属(ActinidiaL ind l.)、荚蒾属(ViburnumL.)和莎草属(CyperusL.);含9种的属有锥属〔Castanopsis(D. Don)Spach〕、石栎属(LithocarpusBl.)、冷水花属(PileaL ind l.)、铁线莲属(ClematisL.)、胡枝子属(LespedezaMichx.)和鼠尾草属(SalviaL.)。在上述被子植物属中,青冈属、冬青属、山矾属、锥属、石栎属、含笑属、樟属、桂樱属、石楠属、蔷薇属和柿属等为该区域森林植被的主要建群属,其他属也多为该区域森林植被的重要组成属。从以上分析中也可看出,崀山丹霞地貌区被子植物种类在属内的分布较分散。
2.1.3 种子植物属的分布区类型 根据种子植物属的现代地理分布特点,并根据吴征镒对中国种子植物属的分布区类型划分理论和方法[13],对崀山丹霞地貌区内的种子植物属的分布区类型进行划分,结果见表2。
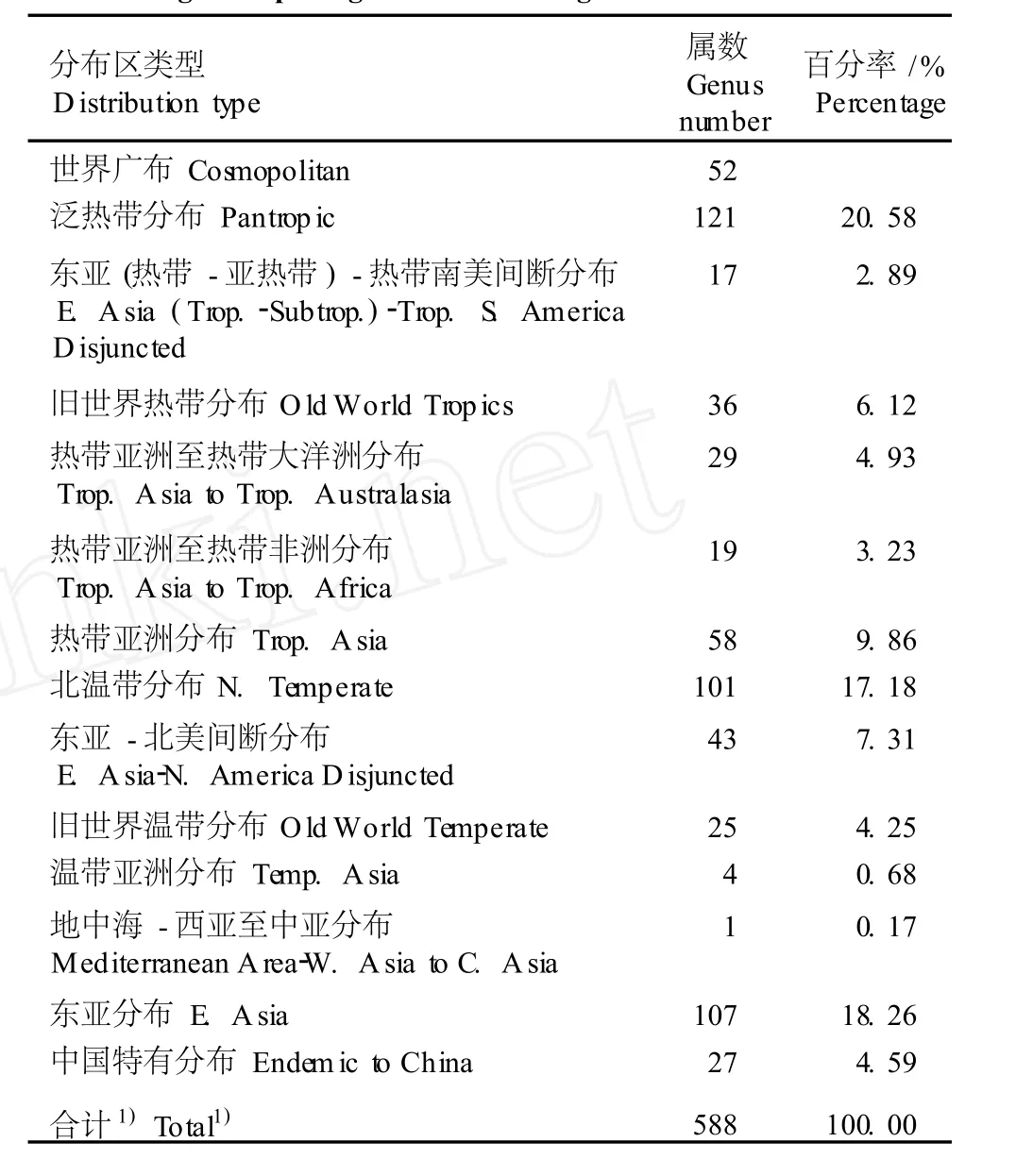
表2 湖南崀山丹霞地貌区种子植物属的分布区类型组成Table2 Com position of genus d istr ibu tion type of seed p lan ts in“Danx ia”geom orpholog ica larea of Langshan Moun ta in in Hu’nan
由表2可以看出,除中亚分布型外,崀山丹霞地貌区内的种子植物属有14个分布区类型。其中,热带分布型属有288属,占该地貌区总属数(不包括世界广布型属)的47.62%,温带分布型属有281属,占该地貌区总属数的47.79%,表现出明显的热带-温带区系分布的特性。
在热带分布型属中,泛热带分布型和热带亚洲分布型占较大的比例。泛热带分布型属中的榕属、冷水花属、花椒属、冬青属、卫矛属、紫金牛属、山矾属、紫珠属(CallicarpaL.)、大戟属(EuphorbiaL.)和薯蓣属等含6种以上,为比较大的属,是该地貌区植物区系的重要成分;其中,冬青属和山矾属所含的种类为群落建群种。泛热带分布型属不仅在该地貌区占较大比例,还包括许多典型成分,如马兜铃属(AristolochiaL.)、马齿苋属(PortulacaL.)、鹿藿属(RhynchosiaLour.)、斑鸠菊属(VernoniaSchreh.)、金合欢属(AcaciaMill.)、巴豆属(CrotonL.)、古柯属(ErythroxylumP. Br.)和三芒草属(AristidaL.)等均有1~2种分布,说明该地貌区与泛热带成分的联系较密切。热带亚洲分布型属包括山桂花属(BennettiodendronMerr.)、山胡椒属、新木姜子属(NeolitseaMerr.)、山茶属、青冈属、含笑属、润楠属(MachilusNees)、蚊母树属(DistyliumSieb.et Zucc.)、南五味子属(KadsuraKaemp f.ex Juss.)、木莲属(ManglietiaBl.)、木荷属(SchimaReinw.)、虎皮楠属(DaphniphyllumBl.)、清风藤属(SabiaColebr.)、唇柱苣苔属(ChiritaBuch.-Ham.ex D.Don)和石斛属(DendrobiumSw.),是该地貌区植物群落的重要组成成分;其中,新木姜子属、山茶属、青冈属、含笑属和润楠属等属所包含的种类为该地貌区植物群落的优势种和次优势种,对丹霞地貌区植物群落的构建起着重要的作用。旧世界热带分布型属、热带亚洲至热带大洋洲分布型属和热带亚洲至热带非洲分布型属的属数在该地貌区总属数中所占比例较小,这些属所包含的种类多为林下、林缘和路边常见种或偶见种,但东亚(热带-亚热带)-热带南美间断分布型属中的木姜子属(LitseaLam.)、楠属(PhoebeNees)、柃木属、山柳属〔Clethra(Gronov.)L.〕和泡花树属(MeliosmaBl.),旧世界热带分布型属中的楝属(MeliaL.)、八角枫属(AlangiumLam.)、海桐花属(PittosporumBanks)、合欢属(AlbiziaDurazz.)、野桐属(MallotusLour.)和楼梯草属,以及热带亚洲至热带大洋洲分布型属中的樟属、栝楼属(TrichosanthesL.)、兰属(CymbidiumSw.)和崖爬藤属〔Tetrastigma(Miq.) Planch.〕等属所包含的种类则为该地貌区植物群落组成成分或群落建群种。
在温带分布型属中,北温带分布型、东亚分布型、东亚-北美间断分布型所占的比例较大。松属(PinusL.)、杨属(PopulusL.)、柳属(SalixL.)、桦木属(BetulaL.)、栎属(QuercusL.)、榆属(UlmusL.)、桑属(MorusL.)、小檗属(BerberisL.)、樱属(CerasusMill.)、花楸属(SorbusL.)、槭属、杜鹃属(RhododendronL.)和荚蒾属等典型的北温带分布型属,以及景天属(SedumL.)、婆婆纳属(VeronicaL.)和乌饭树属(VacciniumL.)等为崀山丹霞地貌区植被的重要成分,其中,松属、栎属和槭属等属包含的种类为崀山丹霞地貌区植物群落建群种。东亚分布型属有107属,包括全型46属、中国-日本变型36属和中国-喜马拉雅变型25属,表现出东部、西部植被成分的过渡交会以及与日本成分广泛联系的特点。东亚-北美间断分布型属所占比例不大,但包含一些具有典型地理分布样式的属,如三白草属(SaururusL.)、鹅掌楸属(LiriodendronL.)、木兰属(MagnoliaL.)、檫木属(SassafrasTrew)、肥皂荚属(GymnocladusLam.)、山核桃属(CaryaNutt.)、香槐属(CladrastisRafin.)、紫藤属(WisteriaNutt.)和枫香树属(LiquidambarL.)等。旧世界温带分布型属、温带亚洲分布型属及地中海-西亚至中亚分布型属与该地貌区植物区系的联系较弱,除少数属的种类参与群落组成外,多数种类为林缘、路边的常见或偶见成分,均不能反映该地貌区植物区系性质。
中国特有分布型属有27属,占总属数的4.59%。包括杜仲属(EucommiaO liv.)、石笔木属(TutcheriaDunn)、香果树属(EmmenopterysO liv.)、青钱柳属(CyclocaryaIljinskaja)、牛鼻栓属(FortuneariaRehd. etWils.)、喜树属(CamptothecaDecne.)、蜡梅属(ChimonanthusL ind l.)、青檀属(PteroceltisMaxim.)、枳属(PoncirusRaf.)、藤山柳属(ClematoclethraMaxim.)、银鹊树属(TapisciaO liv.)、虾须草属(SheareriaS. Moore)、血水草属(EomeconHance.)、半蒴苣苔属、皿果草属(OmphalotrigonotisW.T.Wang)、通脱木属(TetrapanaxK.Koch)、石山苣苔属(PetrocodonHance)、伞花木属(EurycorymbusHand.-Mazz.)、鬼臼属(DysosmaWoodson)、毛药花属(BostrychantheraBenth.)、斜萼草属(LoxocalyxHem sl.)、半枫荷属(SemiliquidambarChang)、杉木属(CunninghamiaR. Br.)、山拐枣属(PoliothyrsisO liv.)、地构 叶 属(SperanskiaBaill.)、匙叶草属(LatoucheaFranch.)、无柱兰属(AmitostigmaSch ltr.)和独花兰属(ChangnieniaS.S.Chien)等,这些属多为一些起源古老的单种属和寡种属(含6种及6种以内的属),反映出该区域植物区系古老起源的特性。其中,香果树属、青檀属、青钱柳属、藤山柳属和杉木属等所包含的种类是崀山丹霞地貌区植物群落的优势种或重要组成种类。以上分析结果说明中国特有分布型属对崀山丹霞地貌区植物区系组成有重要的意义。
2.1.4 古老残遗性与特有成分 一些学者指出,“中国南部是自第三纪以来,甚至白垩纪以来变动不大的古老植物王国”[15]。崀山丹霞地貌区保存了第三纪及第三纪以前的众多古老植物,除卷柏科、石松科(Lycopodiaceae)、木兰科(Magno liaceae)、五味子科(Schisandraceae)、八角科(Illiciaceae)和金缕梅科(Ham am elidaceae)等古老科外,在崀山丹霞地貌区白垩纪就已存在的植被成分有槭属、桦木属、水青冈属(FagusL.)、木兰属、木防己属(CocculusDC.)、山胡椒属、栎属、檫木属和荚蒾属等属;老第三纪就已存在的植被成分有杉木属、松属、三尖杉属(CephalotaxusSieb.et Zucc.)和穗花杉属(AmentotaxusPilger)等属;古新世就已成为中国北方阔叶林成分的榆属、桤木属(AlnusMill.)、鹅耳枥属(CarpinusL.)、柳属和黄杞属(EngelhardiaLesch.ex Bl.)以及亚热带的杨梅属(MyricaL.)、山核桃属、枫香树属和山茱萸属(CornusL.)等属[16]在现有崀山丹霞地貌区植被中成为森林植被的主要成分。崀山丹霞地貌区孑遗植物包括鹅掌楸〔Liriodendronchinense(Hem sl.)Sarg.〕、瘿椒树(TapisciasinensisO liv.)、伞 花 木 〔Eurycorymbus cavaleriei(Levl.)Rehd.et Hand.-Mazz.〕和穗花杉〔Amentotaxusargotaenia(Hance)Pilger〕等种类,有一些古老植物种类还成为单种属或寡种属中的珍稀濒危植物,如:钟萼木(BretschneiderasinensisHem sl.)、杜仲(EucommiaulmoidesO liv.)、鹅掌楸、瘿椒树、青檀(PteroceltistatarinowiiMaxim.)、独花兰(Changnienia amoenaS.S.Chien)和半枫荷(Semiliquidambar cathayensisH.T.Chang)等[17-18],表现出崀山丹霞地貌区植物区系的古老残遗特性。
崀山丹霞地貌区的特殊地质地貌条件与生境的多样性,为该区域特有成分的形成和保存创造了条件。特有成分表现在以下2个方面:有27个中国特有分布型属;有21种新种或新变种的模式或副模式产地位于崀山丹霞地貌区所在县(新宁县)[19]。目前仅发现于新宁县的新种或新变种有13种,其中仅见于崀山丹霞地貌区的种类有新宁毛莨(Ranunculus xinningensisW.T.Wang)和崀山唇柱苣苔(Chirita langshanicaW.T.Wang),是在20多年前发现的新种,经过多年的广泛调查,确认这2个种为仅分布在峎山丹霞地貌区内的特有种,且只生长在崀山丹霞地貌区山体的石壁上,在其他生境条件下无分布,表现出崀山丹霞地貌区的生境狭窄特有现象。
2.2 崀山丹霞地貌区植物区系与舜皇山非丹霞地貌区植物区系的比较分析
为了更进一步说明崀山丹霞地貌区植物区系的特点,选用与崀山丹霞地貌区水平距离约25 km的舜皇山自然保护区的植物区系作对比分析。
2.2.1 区系组成的比较 舜皇山非丹霞地貌区维管植物有792属2232种,其属、种数分别占中国维管植物属、种总数的23.66%和7.72%。其中,蕨类植物有39科86属206种,其属、种数分别占舜皇山非丹霞地貌区维管植物属、种总数的10.86%和9.23%;裸子植物有5科13属19种,其属、种数分别占该区域维管植物属、种总数的1.64%和0.85%;被子植物有693属2007种,其属、种数分别占该区域维管植物属、种总数的87.50%和89.92%。
与舜皇山非丹霞地貌区维管植物相比较,崀山丹霞地貌区维管植物属、种数量分别少了13.01%和34.41%,说明丹霞地貌区维管植物的丰富度低于非丹霞地貌区。崀山丹霞地貌区蕨类植物的科、属、种数分别比舜皇山非丹霞地貌区少35.90%、43.02%和59.22%,其中,缺少了楔叶蕨亚门(Sphenophytina)的木贼目(Equisetales)、真蕨亚门(Filicophytina)厚囊蕨纲(Eusporangiopsida)的瓶尔小草目(Ophioglossales)和原始薄囊蕨纲(Proto lep tospo rangiop sida)的紫萁目(O sm undales)。崀山丹霞地貌区裸子植物的属、种数分别比舜皇山非丹霞地貌区少38.46%和57.89%,被子植物的属、种数分别少8.80%和31.64%。总之,崀山丹霞地貌区蕨类植物科、属、种的数量明显少于舜皇山非丹霞地貌区,裸子植物和被子植物的属和种数量与舜皇山非丹霞地貌区有明显的差异,说明丹霞地貌区干旱的立地条件和孤立的丹霞山地所形成的孤岛[5]对该区域植物区系的形成产生了深刻的影响,出现了特异于非丹霞地貌区的区系组成特征。
2.2.2 种子植物属的分布区类型比较 在舜皇山非丹霞地貌区的种子植物中,热带分布型属有318属,占该区域种子植物总属数(不包括世界广布型属)的48.04%,其中,泛热带分布型、东亚(热带-亚热带)-热带南美间断分布型、旧世界热带分布型、热带亚洲至热带大洋洲分布型、热带亚洲至热带非洲分布型和热带亚洲分布型属的比例分别为19.18%、3.02%、4.83%、3.17%、3.17%和14.65%。温带分布型属有319属,占总属数的48.19%,其中,北温带分布型、东亚-北美间断分布型、旧世界温带分布型、温带亚洲分布型、地中海-西亚至中亚分布型和东亚分布型属的比例分别为19.03%、6.34%、4.83%、0.91%、1.81%和15.26%。中国特有分布型属有25属,占总属数的3.78%。
相比较而言,崀山丹霞地貌区种子植物的热带分布型属和温带分布型属的总比例与舜皇山非丹霞地貌区接近,但2种地貌类型区间各分布区类型所占的比例有一定差异,其中,崀山丹霞地貌区中的热带亚洲至热带大洋洲分布型、旧世界热带分布型、东亚分布型、东亚-北美间断分布型和中国特有分布型属所占的比例明显大于舜皇山非丹霞地貌区,而热带亚洲分布型、温带亚洲分布型以及地中海-西亚至中亚分布型属的比例又明显小于舜皇山非丹霞地貌区,反映出2种地貌类型区种子植物属的各分布区类型间联系的紧密程度存在一定的差异。上述分析结果说明,2种地貌类型区植物区系有着共同的起源属性,但在种子植物属的分布区类型上存在一定的差异。
2.2.3 中国特有成分的比较 舜皇山非丹霞地貌区有中国特有分布型属25属,而模式或副模式产地位于新宁县境内的新种或新变种就有21种,其中的18种在崀山丹霞地貌区也有分布[19],表明舜皇山非丹霞地貌区的植物种类的特有性也是很强的。崀山丹霞地貌区的中国特有成分与舜皇山非丹霞地貌区有一定的差异,其中,崀山丹霞地貌区中国特有分布型属比舜皇山非丹霞地貌区多2属,在崀山丹霞地貌区有分布而在舜皇山非丹霞地貌区无分布的中国特有分布型属有牛鼻山属、皿果草属、半蕨苣苔属、石山苣苔属和无柱兰属等,在舜皇山非丹霞地貌区有分布而在崀山丹霞地貌区无分布的中国特有分布型属有陀螺果属(MelliodendronHand.-Mazz.)、银杉属(CathayaChun et Kuang)和拟单性木兰属(ParakmeriaHu et Cheng),说明2种地貌类型生境对中国特有分布型属的保存有不同的影响。此外,崀山丹霞地貌区特有的新宁毛茛和崀山唇柱苣苔,在舜皇山非丹霞地貌区未曾发现,说明无数孤立的丹霞单体组成的丹霞地貌有利于新种的分化与形成。
3 结 论
湖南崀山丹霞地貌区有野生维管植物185科689属1464种(含变种),其中,蕨类植物25科49属84种、裸子植物5科8属8种、被子植物155科632属1372种,说明该地貌区维管植物多样性较为丰富。在崀山丹霞地貌区分布的被子植物中,含30种以上的大科有12个,共包括233属543种,分别占该地貌区总科数的6.49%、总属数的33.82%、总种数的37.09%;含6种及6种以上的属有52属,共含427种,分别占总属数的7.55%、总种数的29.17%,这些较大的科、属在崀山丹霞地貌区植物区系中占有较重要的位置。
在崀山丹霞地貌区种子植物中,热带分布型属和温带分布型属分别占该地貌区总属数(不包括世界广布型属)的47.62%和47.79%,中国特有分布型属仅占4.59%,具有热带、温带成分交会的特点,其中又以热带成分中的泛热带分布型属和热带亚洲分布型属、温带成分中的北温带分布型属、东亚分布型属和东亚-北美间断分布型属以及中国特有分布型属在崀山丹霞地貌区种子植物区系中占重要地位。崀山丹霞地貌区植物区系具有起源古老、孑遗植物较多、与热带古老成分联系广泛以及特有成分丰富等特性,特有种新宁毛莨和崀山唇柱苣苔的存在,证明崀山丹霞地貌区具有生境狭窄特有现象。
崀山丹霞地貌区与邻近的舜皇山花岗岩中低山地貌区(非丹霞地貌区)比较,均具有植物区系种类相对丰富、热带和温带分布型属所占比例相近等共同特点,但2种地貌区的植物区系组成以及种子植物属的分布区类型存在一定的差异,保存的中国特有成分也不尽相同,说明崀山孤立的丹霞单体组成的丹霞地貌对其植物区系组成产生了深刻影响,且这种特殊的生境也有利于植物新类群的分化与形成以及一些特有成分的延续与保存。崀山丹霞地貌区蕨类植物科、属、种的数量以及裸子植物和被子植物属、种的数量均明显少于非丹霞地貌区,说明丹霞地貌的特殊生境和干旱条件也对植物种类的丰富度产生了很大的影响。
致谢:本论文得到湖南省林业科学院袁正科研究员的指导,特此致谢!
[1]吴志敏,童潜明,陆奇勇,等.龙底沟谷森林生态系统自然保护区地质及景观资料调查研究[M]∥张海浪,刘国平,袁正科.湖南省通道县龙底沟谷森林生态系统自然保护区自然资源研究.长沙:湖南科学技术出版社,2002:45.
[2]中国丹霞申报世界自然遗产协调领导小组.中国丹霞申报世界自然遗产综合文本[R].长沙:中国丹霞申报世界自然遗产协调领导小组,2008:18-33.
[3]袁正科,李星照,彭珍宝,等.崀山丹霞地貌区生物多样性考察与研究综合报告[G]∥中南林业科技大学.中国丹霞申报世界自然遗产提名地——湖南省新宁县崀山生物多样性考察与研究报告.长沙:湖南省新宁县人民政府,中南林业科技大学,2008:2.
[4]叶锦玲.广东丹霞山植被及资源利用的研究[J].仲恺农业技术学院学报,1992(3):37-38.
[5]彭珍宝,旷柏根,夏江林,等.崀山丹霞地貌区野生藤本植物区系与生长特性分析[J].植物资源与环境学报,2009,18(4):58-65.
[6]熊智平,但新球.湖南舜皇山自然保护区综合考察报告[G]∥国家林业局中南林业调查规划设计院.湖南舜皇山自然保护区科学考察集.长沙:国家林业局中南林业调查规划设计院,2006:1-2.
[7]SutherlandWJ.生态学调查方法手册[M].张金屯,译.北京:科学技术文献出版社,1999:127-140.
[8]陈宏伟,于顺利,蔡博峰,等.北京四座楼自然保护区植物区系及植被分析[J].植物资源与环境学报,2006,15(1):62-69.
[9]黄启堂,游水生,陈世品.福建省兰科植物区系分析[J].植物资源与环境学报,2009,18(3):81-87.
[10]祁承经,喻勋林.湖南种子植物总览[M].长沙:湖南科学技术出版社,2001:1-588.
[11]祁承经,林亲众.湖南树木志[M].长沙:湖南科学技术出版社,2000:1-719.
[12]罗仲春,罗毅波,刘小宁,等.湖南舜皇山自然保护区植物区系考察报告[G]∥国家林业局中南林业调查规划设计院.湖南舜皇山自然保护区科学考察报告集.长沙:国家林业局中南林业调查规划设计院,2006:94-109.
[13]吴征镒,周浙昆,孙 航,等.种子植物分布区类型及其起源和分化[M].昆明:云南出版集团公司云南科技出版社,2006:146-451.
[14]谷海燕,李策宏.峨眉山蕨类植物区系的初步研究[J].西北植物学报,2008,28(2):0381-0387.
[15]张灿明,杨秀吉,王佑林,等.龙底沟谷森林生态系统自然保护区植物资源与区系研究[M]∥张海浪,刘国平,袁正科.湖南省通道县龙底沟谷森林生态自然保护区自然资源研究.长沙:湖南科学技术出版社,2004:74.
[16]中国植被编辑委员会.中国植被 [M].北京:科学出版社,1983:6-8.
[17]汪 松,解 焱.中国物种红色名录[M].北京:高等教育出版社,2004:301-400.
[18]张秦伟.秦岭种子植物区系中的珍稀濒危植物[J].植物资源与环境学报,2002,11(3):29-35.
[19]罗仲春,罗毅波.新宁植物 [M].北京:中国林业出版社,2008:1-2.
