基于RAPD标记的叉蕊薯蓣和粉背薯蓣不同居群遗传多样性研究
2010-12-31孙小芹郭建林周义锋白明明杭悦宇
孙小芹,郭建林,周义锋,彭 斌,白明明,杭悦宇
〔江苏省·中国科学院植物研究所(南京中山植物园)江苏省植物迁地保护重点实验室,江苏 南京 210014〕
叉蕊薯蓣(DioscoreacollettiiHook.f.)为薯蓣科(Diosco reaceae)薯蓣属(DioscoreaL.)根状茎组(Sect.Stenophora)植物,为中国特有种之一,主要分布在长江以南各省区。据文献[1-2]记载,叉蕊薯蓣的变种粉背薯蓣〔D.collettiivar.hypoglauca(Palibin)C.T. Ting et al.〕主要分布于安徽、福建、广东北部、广西东北部、河南南部、湖北、湖南、江西、台湾北部及浙江,与原变种的主要区别为药隔分叉程度和蒴果形状不同。叉蕊薯蓣药隔宽度约为花药的1~2倍;粉背薯蓣蒴果两端平截,药隔宽度约为花药的1/2,蒴果上端稍宽,基部狭圆形,顶部与基部同宽[1-2]。Burkill[3]曾根据药隔分叉程度和蒴果形状将叉蕊薯蓣和粉背薯蓣划分为2个种;裴鉴等[4]则认为两者外部形态的差别不严格,故定为原变种与变种。
叉蕊薯蓣是根状茎组中非常独特的具有“3枚雄蕊发育、花药药隔分叉”特征的类群,一些学者依据采自中国各地的标本发表了一些与叉蕊薯蓣相象但似乎又有不同的种类。有些学者认为[5],这些类群分布有交叉,且外部形态的差别不严格,并最终将其归并为1原变种1变种,即原变种叉蕊薯蓣及变种粉背薯蓣。这一认定的依据是:叉蕊薯蓣及粉背薯蓣的分布在四川交叉,且前者向西南、后者向东北直至台湾分布。有的学者还认为[5],两者染色体数有差异,叉蕊薯蓣染色体数为20,粉背薯蓣染色体数为40;但杭悦宇[6]根据对叉蕊薯蓣和粉背薯蓣连续分布区的采集及观察,认为二者在形态上存在着非常明显的连续变化,且叉蕊薯蓣和粉背薯蓣的染色体数均有20和40。基于3个 cpDNA序列的分子系统研究结果则显示[7],叉蕊薯蓣和粉背薯蓣间的亲缘关系极为相近。因此,现有的植物形态学、植物细胞学、植物地理学及分子系统学的研究结果均表明对二者的分类界定应重新考虑。
随着现代生物技术的兴起,各种分子生物学技术和方法为种内变异和亲缘关系的研究提供了新的手段。RAPD(random am p lified po lymorphic DNA,随机扩增多态性DNA)是目前被广泛应用的分子标记之一,它能极其明显地揭示亲缘关系十分相近的个体之间的遗传变异,因此适合于检测种及种下水平的多样性,包括野生植物天然居群遗传结构分析、种质资源评估和栽培植物品种鉴定等[8]。
作者利用RAPD分子标记技术对叉蕊薯蓣及粉背薯蓣分布区内11个居群的遗传多样性进行检测分析,旨在探讨叉蕊薯蓣种内DNA分子水平上的变异与遗传多样性,也为叉蕊薯蓣的分类研究提供一定的参考依据。
1 材料和方法
1.1 材料
叉蕊薯蓣6个居群(包括5个野生居群和1个栽培居群)及粉背薯蓣5个居群的产地见表1。野外调查并采集标本,选取生长状况良好的植株,采集植株中部的健康嫩叶,去除主脉后用体积分数70%乙醇擦去叶片表面灰尘,放入密封袋内用硅胶干燥保存。凭证标本存于江苏省·中国科学院植物研究所,经杭悦宇研究员鉴定。
实验用主要仪器有 PE-9700型 PCR仪(Perkin Elm er公司生产)和WV-BP330型凝胶扫描分析系统(江苏捷达科技发展有限公司生产);使用的10× buffer、dNTPs、Mg2+和TaqDNA聚合酶均购自上海博彩生物科技有限公司,所用引物由上海生工生物工程技术有限公司合成。
1.2 方法
1.2.1 基因组总 DNA的提取 参照 Paterson等[9]的CTAB法提取基因组总DNA,并溶解在灭菌双蒸水中,置于-20℃储存、备用。
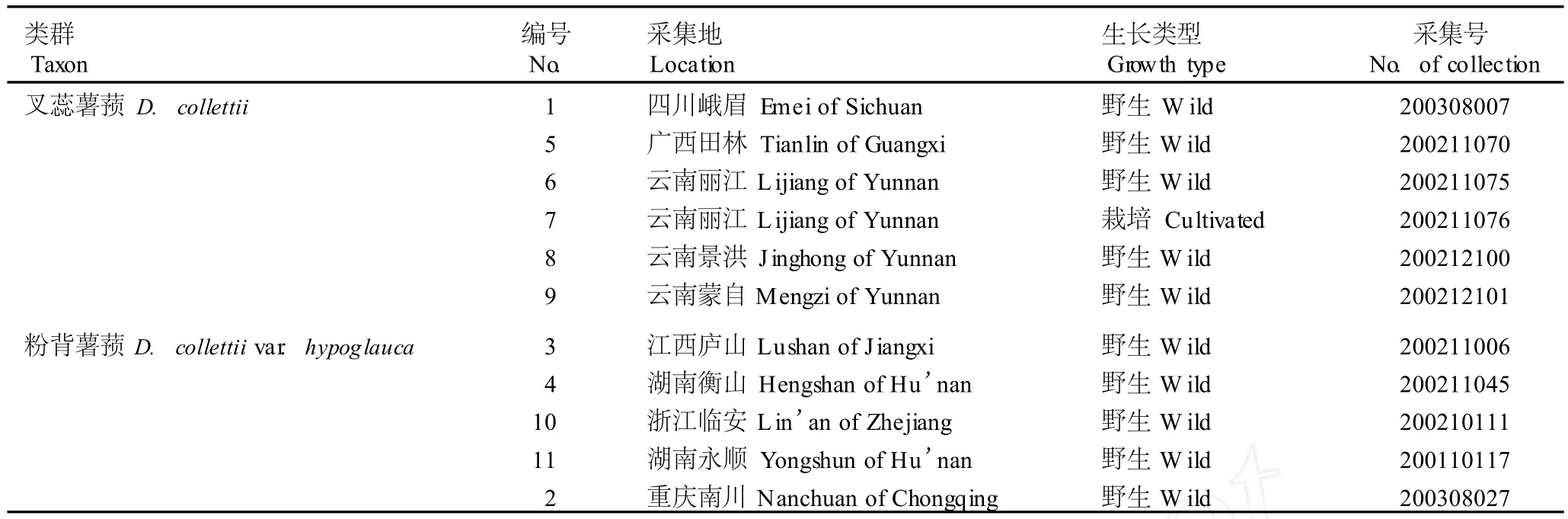
表1 供试叉蕊薯蓣及粉背薯蓣各居群的基本情况Table1 Basic sta tus of tested popu la tion s of D iosco rea co llettii Hook.f.and D.co llettii var.hypog lauca(Pa lib in)C.T.T ing eta l.
1.2.2 引物筛选 采用64个随机引物对叉蕊薯蓣云南丽江居群和粉背薯蓣湖南衡山居群的基因组总DNA进行预扩增,从中筛选出扩增条带清晰、多态性高、重复性好且稳定性强的引物进行全部居群基因组总DNA的RAPD扩增反应。
1.2.3 RAPD-PCR扩增反应条件及扩增结果的检测扩增反应体系总体积为20μL,包含2.0μL10× buffer、0.2 mmo l·L-1dNTPs、2.0 mmol·L-1Mg2+、0.2μmo l·L-1引物、0.4 UTaqDNA聚合酶和20 ng模板DNA,以灭菌双蒸水补足至20μL。
RAPD-PCR扩增反应程序为:95℃预变性3m in;然后于95℃变性30 s、38℃退火40s、72℃延伸1m in,共35个循环反应;最后于72℃保温7m in。扩增产物置于4℃条件下保存。
扩增反应结束后,用质量体积分数0.8%琼脂糖凝胶(含0.5μg·mL-11×EB)对扩增产物进行电泳检测,电泳时间约1 h。电泳结束后用WV-BP330型凝胶扫描分析系统对扩增结果进行观察和拍照。
1.3 数据处理与分析
按照清晰易辨、重复、稳定的原则对扩增谱带进行统计,有条带的记为“1”,同一位置没有条带则记为“0”,据此形成 RAPD表型数据矩阵。统计每个引物扩增出的总条带数及其中的多态性条带数,并计算多态性条带百分率。
应用 POPGENE1.31软件计算有效等位基因数(Ne)、Nei’s基因多样性指数(h)、Shannon多样性指数(I)、基因分化系数(Gst)、基因流(Nm)、遗传相似系数和遗传距离;基于遗传相似系数,采用UPGMA法对11个居群进行聚类分析,构建聚类图。
2 结果和分析
2.1 不同引物 RAPD-PCR扩增结果的比较分析
经过预扩增,从64个随机引物中筛选出扩增条带清晰、多态性高、重复性好且稳定性强的14个寡聚核苷酸引物用于叉蕊薯蓣和粉背薯蓣11个居群基因组总DNA的RAPD-PCR扩增,引物的序列及扩增结果见表2。14个寡聚核苷酸引物共扩增出170条带,平均每个引物扩增出12.1条带,其中多态性条带161条,平均每个引物扩增出的多态性条带数为11.5条,多态性条带百分率达94.71%。其中,引物 a-17、au-04、b-2、b-4、b-5和 b-19扩增出的条带全部为多态性条带,多态性条带百分率均为100.00%。
2.2 叉蕊薯蓣和粉背薯蓣居群的 RAPD-PCR扩增结果及遗传变异分析
由叉蕊薯蓣和粉背薯蓣各居群的 RAPD-PCR扩增结果(表3)可见,5个粉背薯蓣居群的多态性条带百分率变化幅度相对较大,为81.25%~89.29%,而6个叉蕊薯蓣居群的多态性条带百分率变化幅度相对较小,为82.00%~84.21%。在参试的11个居群中,粉背薯蓣江西庐山居群扩增出的多态性条带最多,达75条,多态性条带百分率也最高,达89.29%;粉背薯蓣浙江临安居群扩增出的多态性条带最少,仅39条,多态性条带百分率也最低,仅81.25%。
尽管叉蕊薯蓣和粉背薯蓣的外部形态高度相似,但从不同居群多态性条带百分率的变化幅度可以看出粉背薯蓣遗传多样性更丰富,间接显示区域比形态更易造成二者的遗传分化,同属同组狭域分布的盾叶薯蓣(D.zingiberensisC.H.Wright)居群间的多态性条带百分率较低也证明了这一点[10]。
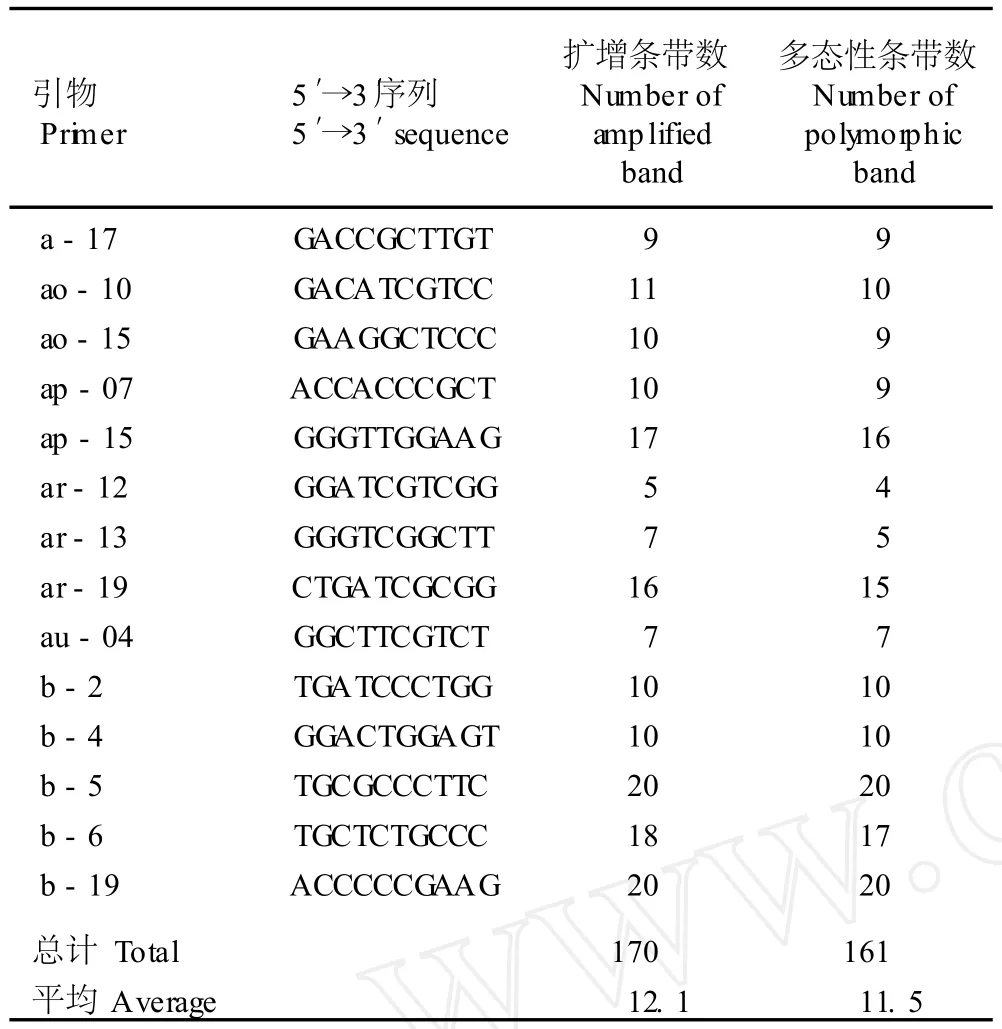
表2 用于叉蕊薯蓣和粉背薯蓣基因组总DNA RAPD-PCR的随机引物碱基序列及扩增结果Table2 Base sequences of random pr im ers used for RAPD-PCR of genom ic tota lDNA of D iosco rea co llettii Hook.f.and D.co llettii var. hypog lauca(Pa lib in)C.T.T ing eta l.and itsam p lified resu lts

表3 叉蕊薯蓣和粉背薯蓣各居群基因组总DNA的 RAPD-PCR扩增结果Table3 RAPD-PCR am p lified resu lts of genom ic tota l DNA of d ifferen t popu la tion s of D iosco rea co llettii Hook.f.and D.co llettii var.hypog lauca(Pa lib in)C.T.T ing eta l.
统计和计算结果表明,粉背薯蓣各居群间的有效等位基因数(Ne)、Nei’s基因多样性指数(h)和Shannon多样性指数(I)分别为1.3685、0.2384和0.3763;叉蕊薯蓣各居群间的Ne、h和I分别为1.3311、0.1972和0.2983。叉蕊薯蓣与粉背薯蓣的基因分化系数(Gst)为0.1228,即在总的变异中,有12.28%的变异存在于这一个类群间,87.72%的变异存在于各自的居群间,表明叉蕊薯蓣和粉背薯蓣作为原变种和变种的相对独立性。同时,也有研究表明单子叶植物近缘种的基因分化系数平均值为0.231,表示有76.9%的变异存在于种内居群间[11]。结合上述研究结果可见,叉蕊薯蓣和粉背薯蓣的遗传变异符合“种内遗传变异大于种间遗传变异”的普遍性规律。
叉蕊薯蓣和粉背薯蓣之间的基因流(Nm)为3.5707,远远高于盾叶薯蓣的Nm(0.9641)[10]及薯蓣科近缘植物延龄草(TrilliumtschonoskiiMaxim.)的Nm(0.4002)[12],表明叉蕊薯蓣和粉背薯蓣之间存在非常多的基因交流。
2.3 叉蕊薯蓣和粉背薯蓣各居群的遗传距离及聚类分析
参试的叉蕊薯蓣6个居群和粉背薯蓣5个居群间的遗传距离和遗传相似系数见表4。由表4可见,粉背薯蓣江西庐山居群和叉蕊薯蓣云南丽江居群的遗传距离最远,达0.6931;叉蕊薯蓣云南蒙自居群和云南景洪居群间的遗传距离最近,仅0.2194。叉蕊薯蓣与粉背薯蓣间的遗传相似系数高达0.9223,但遗传距离仅为0.0809。
基于叉蕊薯蓣和粉背薯蓣居群间的遗传相似系数(表4),采用UPGMA聚类分析方法获得的参试11个居群的聚类图见图1。由图1可见,参试的6个叉蕊薯蓣居群和5个粉背薯蓣居群可分成4组,即所有的叉蕊薯蓣居群和粉背薯蓣浙江临安居群聚为第1组,粉背薯蓣重庆南川居群和湖南衡山居群聚为第2组,粉背薯蓣湖南永顺居群和江西庐山居群则各自单独成组,且粉背薯蓣江西庐山居群与其他居群的亲缘关系最远。
聚类分析结果表明,叉蕊薯蓣及粉背薯蓣11个居群的分组情况是以类群为主要依据的,而在各类群内部则基本以区域分布为分组依据,惟一例外的是粉背薯蓣浙江临安居群与叉蕊薯蓣的6个居群同组,虽然和绝大多数叉蕊薯蓣居群的亲缘关系较远,但却较叉蕊薯蓣四川峨眉居群与绝大多数叉蕊薯蓣居群的亲缘关系更近。
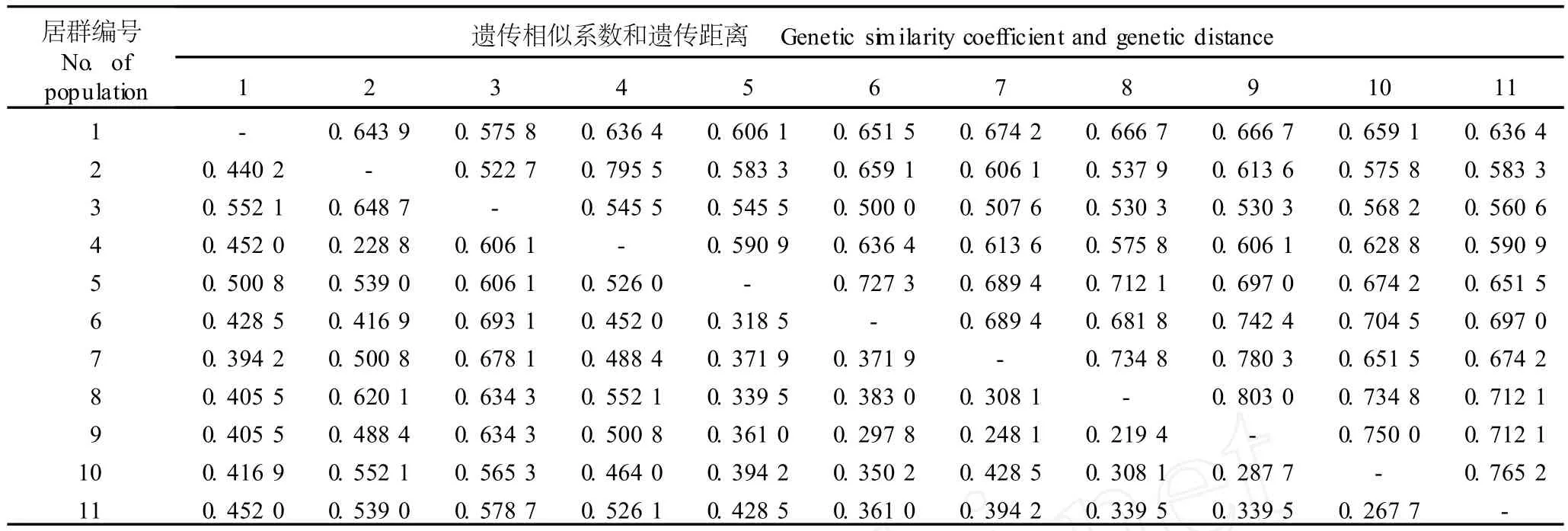
表4 叉蕊薯蓣和粉背薯蓣各居群间的遗传相似系数和遗传距离1)Table4 Genetic sim ilar ity coefficien t and genetic d istance am ong d ifferen t popu la tion s of D iosco rea co llettii Hook.f.and D.co llettii var. hypog lauca(Pa lib in)C.T.T ing eta l.1)

图1 基于遗传相似系数的叉蕊薯蓣和粉背薯蓣不同居群的 UPGMA聚类图Fig.1 UPGMA dendrogram of d ifferen t popu la tion s of D iosco rea co llettii Hook.f.and D.co llettii var.hypog lauca(Pa lib in)C.T. T ing eta l.based on genetic sim ilar ity coefficien t
3 讨 论
叉蕊薯蓣是 Hooker JD在1892年依据采自缅甸掸邦地区的标本订名发表的。此后,一些学者依据采自中国各地的标本发表了一些主要特征与叉蕊薯蓣相似的新种,如D.hypoglaucaPalibin(福建葛岭)、黑叶薯蓣D.nigrescensKnuth(云南思茅)、D.morseiPrain et Burkill(江西庐山牯岭)、D.oeneaPrain et Burkill(四川城口)、D.kelungensisHayata(台湾基隆)、D.tashiroiHayata(台湾)和D.undulateKnuth(重庆南川)等[13],但随着研究的不断深入,学者们认为这些种可归并为数种[14]、2种[5]、1种[15-16]或1正种及1变种[1]。裴鉴等[4]认为,除黑叶薯蓣外,这些种类全部都在粉背薯蓣的分布区域内,因而将它们都归并入叉蕊薯蓣或粉背薯蓣,并将叉蕊薯蓣定为原变种、粉背薯蓣定为变种。尽管叉蕊薯蓣分布区较窄,但是形态分化却较大,如叉蕊薯蓣的云南丽江居群叶片大、草质程度高;云南蒙自居群果实少、透明,全茎、花梗及果实均为鲜艳的紫红色等等,这些都充分证明了其分布地为强烈分化的起源地。然而,不同分布区的粉背薯蓣形态基本一致,较少出现分化现象。
鉴于薯蓣属植物为雌雄异株,本研究结果又表明叉蕊薯蓣和粉背薯蓣具有很高的遗传多样性(多态性条带百分率在81%以上)和基因流(3.5707),在一定程度上说明野生状态下它们的繁殖过程主要通过有性繁殖的方式来完成。叉蕊薯蓣和粉背薯蓣所在的薯蓣属根状茎组植物的传粉方式有虫媒[17]和风媒[18]等多种途径,而且雌雄花完全开放且不具有零余子,表明无论采用何种传粉方式,根状茎组植物都通过有性繁殖途径繁衍后代[18]。尽管薯蓣属根状茎组植物的根茎具有无性繁殖能力,但是在野外状况下,1个根状茎通常只生长出1支缠绕茎,没有出现由于根状茎逐年增生而在1个基株上生长出多个单株的情况[18-19],这也证明了薯蓣属根状茎组植物在野生状况下主要的繁殖方式为有性繁殖。
在聚类图上,粉背薯蓣浙江临安居群和叉蕊薯蓣聚合在一起;从形态上看,粉背薯蓣浙江临安居群和粉背薯蓣其他居群有所差别,其花、茎和果实的颜色为绿色中带有紫红色,这一特征和叉蕊薯蓣云南蒙自居群相似。推测出现这种现象的原因有2个:①作为原变种和变种的叉蕊薯蓣和粉背薯蓣可能还存在着变种水平上的分化不完全;②叉蕊薯蓣和粉背薯蓣本身即为形态连续变化的1个类群。基于 cpDNA的分子系统学研究结果表明,叉蕊薯蓣和粉背薯蓣的trnL-F、rbcL和matK序列碱基有10个以内的差异,在系统发生树上两者合为一支,支持率为60%[7],也证明了叉蕊薯蓣和粉背薯蓣存在着变种水平的分化不完全。但叉蕊薯蓣和粉背薯蓣在地下茎解剖特征[20]、淀粉粒形态[21]、叶脉序结构以及叶表皮解剖特征[6]上又存在着明显的差异,如:叉蕊薯蓣具导管而粉背薯蓣具管胞;叉蕊薯蓣地下茎淀粉粒偶有由2~3小粒淀粉粒组成的复粒且长度为14~22μm,而粉背薯蓣地下茎淀粉粒无复粒且长度为25~47μm;叉蕊薯蓣叶脉的末次脉边缘结环状而粉背薯蓣不结环;粉背薯蓣叶表皮存在二叉表皮毛而叉蕊薯蓣没有,这些不同的特征均表明粉背薯蓣有异于叉蕊薯蓣。综合以上结果认为,叉蕊薯蓣和粉背薯蓣为原变种和变种的关系,两者间还存在着变种水平上的分化不完全。
[1]中国科学院中国植物志编辑委员会.中国植物志:第十六卷第一分册[M].北京:科学出版社,1985:54-120.
[2]D ing Z Z,GilbertMG.Flora of China:Vo l.24[M].Beijing: Science Press,2000:276-296.
[3]Burkill FLSI H.The o rganography and the evo lution of D iosco reaceae,the fam ily of the Yam s[J].Journal of the L innean Society of London:Botany,1960,56:319-412.
[4]裴 鉴,丁志遵,秦慧贞,等.中国薯蓣属根茎组系统分类的初步研究[J].植物分类学报,1979,17(3):61-72.
[5]江苏省植物研究所薯蓣课题研究组.中国薯蓣属根状茎组植物的分类和染色体数的研究[J].植物分类学报,1976,14(1):65-72.
[6]杭悦宇.中药萆薢的原植物、鉴定及质量评价研究[D].南京:中国药科大学生药学院,2007:48-55.
[7]Gao X,Zhu Y P,Wu BC,et al.Phylogeny ofDioscoreaSect.Stenophorabased on chlorop lastmatK,rbcLandtrnL-Fsequences [J].Journal of Systematics and Evolution,2008,46(3):315-321.
[8]邹喻苹,葛 颂,王晓东.系统与进化植物学中的分子标记[M].北京:科学出版社,2001.
[9]Paterson A H,Brubaker C L,Wendel J F.A rap id method for extraction of cotton(Gossypiumspp.)genom ic DNA suitable for RFLP or PCR analysis[J].Plant Molecular Biology Reporter,1993,11(2):122-127.
[10]黄春洪,杭悦宇,周义锋,等.我国盾叶薯蓣居群遗传结构分析[J].云南植物研究,2003,25(6):641-647.
[11]Ham rick JL.Isozymesand the analysis of genetic structure in p lant populations[M]∥Soltis DE,So ltis P S.Isozymes in Plant Bio logy.London:Chapman and Hall,1990:87-105.
[12]李 群,肖 猛,郭 亮,等.四川省珍稀濒危植物延龄草遗传多样性分析[J].北京林业大学学报,2005,27(4):1-6.
[13]万金荣,丁志遵,周太炎.叉蕊薯蓣类群分类学的初步研究[M]∥南京中山植物园研究论文集编辑组.南京中山植物园研究论文集:1990.南京:江苏科学技术出版社,1991:41-45.
[14]Knuth R.D ioscoreaceae[M]∥Engler A.Das Pflanzenreich.Leipzig:Verlag vonWilhelm Engelmann,1924:252-254,315.
[15]中国科学院昆明植物研究所.云南植物志:第三卷[M].北京:科学出版社,1983:717-719.
[16]中国科学院昆明植物研究所.云南种子植物名录:下册[M].云南:云南人民出版社,1984:1981-1982.
[17]赵亚美,杭悦宇,周义峰,等.柴黄姜的传粉生物学特性[J].植物资源与环境学报,2008,17(2):15-21.
[18]赵亚美.薯蓣属(DioscoreaL.)植物传粉生物学研究[D].南京:江苏省·中国科学院植物研究所,2008:29-34.
[19]王筱璐,吴宝成,周义峰,等.野生及栽培盾叶薯蓣(Dioscorea zingiberensis)的性别特征观察 [J].植物资源与环境学报,2010,19(2):15-20.
[20]杭悦宇,秦慧贞,徐珞珊,等.薯蓣属根状茎的组织结构和萆薢类鉴别[J].西北植物学报,2006,26(12):2423-2429.
[21]杭悦宇,徐珞珊,史德荣,等.中国薯蓣属植物地下茎淀粉粒形态特征及其分类学意义[J].植物资源与环境学报,2006,15(4):1-8.
