镧对大鼠海马CA1区环磷酸腺苷应答元件结合蛋白磷酸化的影响
2010-11-12杨敬华巫生文刘秋芳张立丰
杨敬华,巫生文,刘秋芳,张立丰,齐 鸣,鲁 帅,蔡 原
(中国医科大学公共卫生学院卫生毒理学教研室,辽宁沈阳 110001)
近十几年来,稀土元素在我国的应用涉及农业、畜牧业和水产养殖等多个领域,越来越多的稀土元素进入环境,并通过食物链进入人体。稀土元素对生物体具有毒物兴奋效应作用,即低水平的稀土元素能促进机体的生长,而高水平的稀土元素对机体具有毒性作用[1]。人群调查显示,稀土矿区儿童的智商均数和学习记忆等认知能力明显低下[2]。镧属于自然界中含量较多的轻稀土元素。动物实验显示,镧可以影响神经行为发育以及学习记忆能力[3-4],但其机制仍未完全阐明。海马是脑内与学习记忆有关的重要结构,即刻早期基因c-fos和c-jun与学习记忆关系密切[5]。本实验室前期的研究已发现,镧损害学习记忆的机制与海马CA1区c-fos表达水平降低有关[6]。cAMP应答元件结合蛋白(cAMP response element binding protein,CREB)是诱导即刻早期基因转录的关键性因素,在学习记忆中发挥重要作用[7]。CREB的Ser133位点磷酸化是其活化形式,钙调蛋白依赖性蛋白激酶Ⅳ(calmodulin dependent protein kinaseⅣ,CaMKⅣ)是磷酸化CREB Ser133位点的重要蛋白激酶[8]。因此,观察氯化镧(lanthanum chloride,LaCl3)对大鼠海马CA1区CaM活性和CaMKⅣ,CREB磷酸化以及c-jun基因转录和蛋白表达的影响,研究镧的毒性及其作用机制。
1 材料与方法
1.1 试剂
LaCl3,分析纯(国药沈阳化学试剂有限公司),蛋白定量试剂盒(普利来基因技术有限公司),RTPCR试剂盒(TaKaRa BIO),磷酸二酯酶,环磷腺苷和蝮蛇蛇毒(Sigma),兔抗大鼠p-CaMKⅣ抗体,兔抗大鼠p-CREB抗体,兔抗大鼠c-jun抗体和兔抗大鼠GAPDH抗体(Santa Cruz)。
1.2 仪器
GRADIENT96型PCR仪(德国,Biometra公司)、X-7电感耦合等离子体质谱仪(美国,Thermo Elemental公司)、CS150GXL型低温高速离心机(日本,Hitachi公司)、JY96-IIN型超声波细胞破碎机(中国,宁波新芝生物科技股份有限公司)、HH42型快速恒温数显水箱(中国,常州国华电器有限公司)、PAC300水平电泳仪(美国,Bio-Rad Power公司)、DYY-6B型垂直电泳仪(中国,北京六一仪器厂)和IMAGER5500凝胶图像分析系统(美国,Alpha公司)。
1.3 动物分组与染毒
清洁级 Wistar大鼠〔证号:SYXK(辽),2003-0013〕由中国医科大学实验动物中心提供,60只,体重250~270 g,雌雄比例2∶1。正式实验前饲养7 d,实验动物室温度17~23℃,相应湿度45% ~55%,动物饲料由实验动物中心提供。将40只雌性Wistar大鼠随机分为对照组、LaCl30.25%、0.5% 和1.0%染毒组,然后将雌性大鼠与雄性大鼠按2∶1同笼进行交配,次日晨发现阴栓或阴道分泌物镜检有精子者定为受孕,记为妊娠第0天。对照组孕鼠饮用蒸馏水。镧染毒组仔鼠在断乳前经由吸吮母乳染镧,断乳后则通过自行饮用LaCl30.25%,0.5%和1.0%蒸馏水溶液染镧1个月,然后进行各项指标测定。每项指标均从每组随机取8只仔鼠进行检测。
1.4 电感耦合等离子体质谱法[9]检测仔鼠海马CA1区镧含量
仔鼠麻醉后,用生理盐水全身灌流以消除血液的影响。取海马CA1区组织,置于特氟纶(Teflon)管内,加入65%硝酸2 ml和H2O21 ml,用特氟纶塞密封后置于钢罐中。钢罐密封后在180℃加热4 h以消化海马组织样品,然后用2%(V/V)硝酸稀释已消化的组织样品至10 ml,采用电感耦合等离子体质谱仪测定海马CA1区组织样品中的镧含量。仪器工作参数:入射功率:1200 W;反向功率:3 W;雾化气流速:0.78 L·min-1;辅助气流速:0.70 L·min-1;冷却气流速:13.0 L·min-1;样品提升量:1.2 ml·min-1;质量采集方式:脉冲;数据采集方式:跳峰;停留时间:10 ms;扫描次数:50;分辨率:l25;每个质量通道数:3。在上述工作条件下,镧标准系列溶液和组织样品液依次进入ICP-MS后,以镧标准溶液浓度和对应信号值,绘制标准曲线,在标准曲线上查出组织样品液中镧浓度。然后按下式计算镧含量:组织样品中镧含量(μg·g-1组织)=组织样品液镧浓度(mg·L-1)×组织样品液总体积(ml)×组织样品的质量(g)-1。
1.5 磷酸二酯酶法[10]检测仔鼠海马CA1区CaM活性
取海马CA1区组织,充分匀浆,超声波破碎细胞,20 000×g离心15 min,取上清,加入含PDE的反应缓冲液,再加入20 mmol·L-1的环磷腺苷以启动反应,转入30℃水浴下反应30 min。反应到时后转移至沸水浴中终止反应,然后立即移至冰浴中冷却,再加入蝮蛇蛇毒1 g·L-1,于30℃水浴中反应,加入55%TCA终止反应。以20 000×g离心15 min,取上清液,加入定磷试剂,在850 nm处测定吸光度值(A)。以KH2PO4标准系列溶液浓度和对应A值,绘制标准曲线,在标准曲线上查出组织样品液中磷浓度。CaM的1个活性单位(U)是指在上述反应条件下,每分钟由底物生成1μmol磷。按下式计算CaM活性:组织样品中CaM活性(U·g-1蛋白)=样品液磷浓度(mmol·L-1)×样品液总体积(ml)×组织蛋白含量(g)-1×时间(min)-1。
1.6 Western印迹法检测仔鼠海马 CA1区p-CaMKIV,p-CREB和c-jun蛋白含量
取海马CA1区组织,提取蛋白,采用酚试剂法进行蛋白定量。取总量相同的蛋白,在10%SDS-聚丙烯酰胺中电泳,然后将蛋白转移至硝酸纤维素膜上,将膜于4℃、5%脱脂奶粉中封闭2 h,TTBS洗3次,兔抗大鼠 p-CaMKⅣ(1∶100)、p-CREB 抗体(1∶200)、c-jun 抗 体 (1∶200)或 GAPDH 抗 体(1∶1000)中4℃孵育过夜,TTBS洗3次,室温下二抗中孵育2 h。应用ECL显示反应产物,曝光,显影,定影。采用图像分析系统进行蛋白表达水平分析,兔抗GAPDH抗体作为内参校正上述蛋白的相对表达水平。积分吸光度 A(integrated absorbance,IA)表示海马中蛋白的表达水平。
1.7 RT-PCR法检测仔鼠海马 CA1区 c-jun mRNA水平
取海马CA1区组织,提取总RNA,进行RNA定量与纯度鉴定。用TaKaRa RT-PCR Kit(AMV)的二步法进行 RT-PCR。c-jun引物序列:上游5'-AGCAATGGGCACATCACC-3',下游5'-TCTGCGGCTCTTCCTTCA-3',PCR产物长度451 bp;β肌动蛋白引物序列:上游 5'-TTTTGTGCCTTGATAGTTCGC-3',下游5'-GAGTCCTTCTGACCCATACCC-3',PCR 产物长度264 bp。所得产物行琼脂糖凝胶电泳,凝胶图像分析系统进行灰度扫描,以c-jun条带灰度值与β肌动蛋白条带灰度值的比值表示c-jun mRNA的表达水平。
1.8 统计学分析
2 结果
2.1 镧染毒仔鼠海马CA1区镧含量
与正常对照组相比,各染毒组仔鼠海马CA1区镧含量显著增高(P<0.05),并与镧染毒剂量呈正相关,相关系数 r=0.989(P <0.05)(表1)。

表1 染毒仔鼠海马CA1区镧含量Tab.1 Lanthanum contents in the hippocampal CA1 area of rat pups after lanthanum exposure
2.2 镧对仔鼠海马CA1区钙调蛋白活性的影响
表2结果显示,与正常对照组相比,镧染毒使仔鼠海马CA1区钙调蛋白活性降低(P<0.05),并随着染毒剂量的增加而逐渐降低,LaCl30.25%,0.5%和1.0%染毒组仔鼠海马CA1区钙调蛋白活性分别降低至正常对照组的 80.7%,59.9%和 30.6%(P<0.05),说明镧染毒以剂量依赖性方式下调钙调蛋白活性。
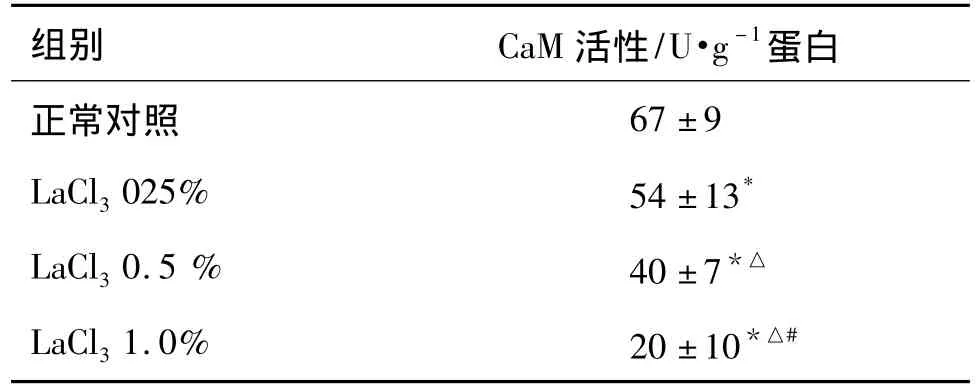
表2 镧对仔鼠海马CA1区钙调蛋白(CaM)活性的影响Tab.2 Effect of lanthanum on calmodulin(CaM)activity in the hippocampal CA1 area of rat pups
2.3 镧对仔鼠海马CA1区p-CaMKⅣ蛋白表达的影响
图1结果显示,与正常对照组比,LaCl30.25%,0.5%和1.0%染毒使仔鼠海马CA1区p-CaMKⅣ蛋白表达降低,分别降低至正常对照水平的83.0%,57.4%和27.7%(P <0.05),说明镧染毒以剂量依赖性方式下调p-CaMKⅣ蛋白表达水平。
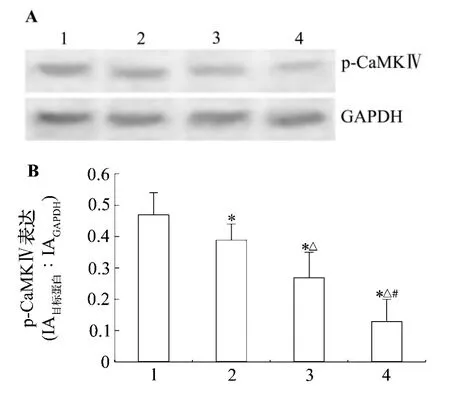
图1 镧对仔鼠海马CA1区磷酸化钙调蛋白依赖性蛋白激酶Ⅳ(pCaMKⅣ)蛋白表达的影响.动物处理见表1.1:正常对照组;2-4:LaCl3 0.25%,0.5%和1.0%组.±s,n=8.*P<0.05,与正常对照组比较;△P <0.05,与 LaCl3 0.25% 组比较;#P <0.05,与 LaCl3 0.5%组比较.Fig.1 Effect of lanthanum on phosphorylated-calmodulin dependent protein kinaseⅣ(p-CaMKⅣ)in the hippocampal CA1 area of rat pups.
2.4 镧对仔鼠海马CA1区p-CREB蛋白表达的影响
图2 结果显示,与正常对照组比,LaCl30.25%,0.5%,1.0%染毒使仔鼠海马CA1区p-CREB蛋白表达降低,分别降低了 24.4%,36.6%和 73.2%(P<0.05),说明镧染毒与 p-CREB蛋白表达下降之间具有一定的剂量-反应关系。
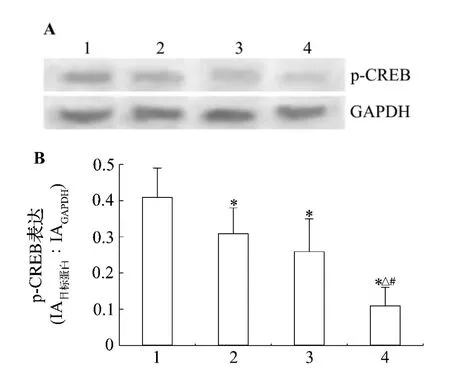
图2 镧对仔鼠海马CA1区磷酸化环磷酸腺苷应答元件结合蛋白(p-CREB)蛋白表达的影响.动物处理见表1.1:正常对照组;2-4:LaCl3 0.25%,0.5%和1.0%组.±s,n=8.*P<0.05,与正常对照组比较;△P <0.05,与 LaCl3 0.25% 组比较;#P <0.05,与 LaCl3 0.5%组比较.Fig.2 Effect of lanthanum on phosphorylated-cAMP response element binding protein(p-CREB)in the hippocampal CA1 area of rat pups.
2.5 镧对仔鼠海马CA1区c-jun mRNA表达的影响
图3结果显示,与正常对照组比,LaCl30.25%,0.5%和1.0%染毒使仔鼠海马CA1区c-jun mRNA表达降低,分别降低了14.8%,27.2%和76.5%(P <0.05),说明镧与 c-jun mRNA 表达下降之间具有一定的剂量-效应关系。
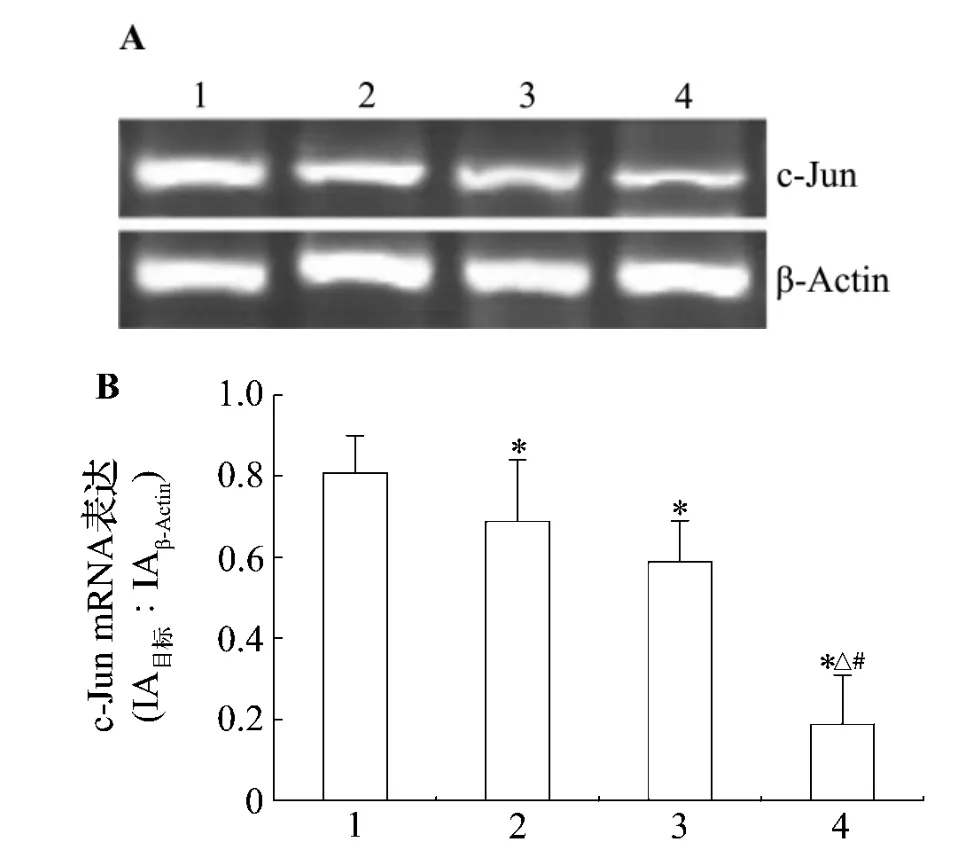
图3 镧对仔鼠海马CA1区c-jun mRNA表达的影响.动物处理见表1.1:正常对照组;2-4:LaCl3 0.25%,0.5%和1.0%组.x¯±s,n=8.*P<0.05,与正常对照组比较;△P<0.05,与LaCl3 0.25%组比较;#P <0.05,与 LaCl3 0.5%组比较.Fig.3 Effect of lanthanum on c-jun mRNA in the hippocampal CA1 area of rat pups.
2.6 镧对仔鼠海马CA1区c-jun蛋白表达的影响

图4 镧对仔鼠海马CA1区c-jun蛋白表达的影响.动物处理见表1.1:正常对照组;2-4:LaCl3 0.25%,0.5%和1.0%组.x¯±s,n=8.*P<0.05,与正常对照组比较;△P<0.05,与LaCl3 0.25%组比较;#P <0.05,与 LaCl3 0.5%组比较.Fig.4 Effect of lanthanum on c-jun protein in the hippocampal CA1 area of rat pups.
图4 结果显示,与正常对照组比,LaCl30.25%,0.5%,1.0%染毒使仔鼠海马CA1区c-jun蛋白表达降低,分别降低了36.1%,45.9%和83.6%(P <0.05),说明镧染毒与c-jun蛋白表达下降之间具有一定的剂量-反应关系。
3 讨论
有研究表明,REEs可进入脑组织,并在脑组织内长期蓄积,其中镧在脑组织中的蓄积性较强[11]。Feng等[12]在大鼠4周 ~6月龄期间 ig给予 LaCl30.1,2 和 40 mg·kg-1染毒,结果发现 LaCl340 mg·kg-1染毒组大鼠的脑组织内明显蓄积,大脑皮质、海马和小脑中镧含量分别达到0.039,0.014和0.015μg·g-1组织。本研究显示,出生后至断乳后 1个月期间LaCl3暴露造成镧在大鼠海马CA1区明显蓄积,镧在海马CA1区的蓄积量与染镧剂量呈明显正相关。任丽宝等[13]对镧在大鼠脑组织中吸收规律和蓄积特点的研究结果也显示,大鼠脑组织中镧含量随其摄入量增加而逐渐增高,存在一定的量效关系。海马是脑内与学习记忆有关的重要结构和关键部位,海马包括CA1,CA2和CA3区和齿状回区域,其中CA1区和学习记忆关系更为密切。因此,本研究中LaCl3大于0.25%暴露造成镧在海马CA1区蓄积,这可能是LaCl3影响中枢神经系统(尤其是海马)结构和功能的基础。
CaM是一个广泛存在于真核细胞生物中的多功能蛋白,它广泛分布于脑组织中。CaM在细胞内有两种存在形式,一种是非活性CaM,以游离状态存在于细胞的胞浆里;一种是活性CaM,与Ca2+和酶/蛋白结合。当CaM与Ca2+结合时,形成Ca2+-CaM复合物,CaM构象发生改变,暴露出肽链表面的疏水基团,保证CaM与靶酶/蛋白的结合完全匹配[14]。CaM在Ca2+参与的细胞信号转导过程中起关键作用,而且CaM直接参与长时程增强的形成[15]。在本研究中,染镧造成仔鼠海马 CA1区CaM活性显著降低,可见染镧造成仔鼠海马中具有生物活性的CaM水平下降,其原因可能是由于CaM是细胞内La3+作用的靶点之一,La3+与Ca2+竞争结合于CaM,La3+与CaM的结合力强于Ca2+与CaM的结合力,影响 Ca2+与 CaM 的正常结合[16-18],从而造成海马中具有生物活性的CaM水平降低。随着LaCl3染毒剂量的增加,仔鼠海马CaM活性进一步下降,其原因可能是更多的La3+与Ca2+竞争结合于CaM,造成具有生物活性的CaM水平进一步降低。
CaMK包括 CaMKⅠ,CaMKⅡ,CaMKⅢ和CaMKⅣ,其活化有赖于CaM。其中CaMKⅣ又称做CaMK Gr,主要在神经元的细胞核中表达,它是磷酸化转录因子CREB的重要蛋白激酶,在突触可塑性、学习记忆过程中发挥重要作用[19]。磷酸化对于CaMKⅣ的激活是必需的,即磷酸化的CaMKⅣ是其活化形式。在本研究中,染镧组仔鼠海马CA1区p-CaMKⅣ蛋白表达水平显著低于对照水平,可见在LaCl3的作用下,由于仔鼠海马CA1区CaM活性显著降低,激活CaMKⅣ的能力下降,造成仔鼠海马CA1区活化的CaMKⅣ(即p-CaMKⅣ)表达水平显著减少,CaMKⅣ的生物学作用受到削弱。
CREB属于含亮氨酸拉链的转录因子家族成员,是细胞内多种信号通路的一种关键成分,具有调节包括学习记忆在内的广泛的生物学功能。研究表明,CREB依赖性转录是学习记忆和可塑性形成过程所必需的[20]。在本研究中,镧染毒组仔鼠海马CA1区p-CREB的表达水平显著低于对照水平,可见染镧造成仔鼠海马CA1区CREB的磷酸化水平显著下降。c-jun基因是即刻早期基因家族成员,CREB的Ser133位点磷酸化是触发c-jun转录的关键所在。研究显示,c-jun与c-fos基因的表达产物结合成复合物,作用于异源二聚体激活蛋白-1结合位点,激活编码突触成分、影响突触传递的基因,从而较长时间地改变神经元的结构和功能,影响突触可塑性[21-22]。本研究显示,镧染毒组仔鼠海马CA1区c-jun mRNA和蛋白表达水平明显低于正常对照组水平,c-jun蛋白表达水平的变化趋势与其基因转录水平的变化一致,而且海马CA1区c-jun mRNA和蛋白表达水平和染毒剂量之间具有一定的剂量-效应关系,说明镧染毒造成海马CA1区c-jun基因转录和蛋白表达水平降低,损害突触可塑性,因而损害大鼠的学习记忆能力。
本研究表明镧损害学习记忆能力的机制可能与镧造成海马CA1区CaM活性和CaMKⅣ、CREB磷酸化以及c-jun基因转录和蛋白表达水平下降有关,但镧损害学习记忆能力的确切机制尚需进一步探讨。
[1]徐厚恩.中国资源环境与健康研究进展[M].北京:北京医科大学出版社,2004:43-64.
[2]范广勤,袁兆康,郑辉列,刘志刚.儿童稀土暴露的健康效应研究[J].卫生研究,2004,33(1):23-28.
[3]Feng L,Xiao H,He X,Li Z,Li F,Liu N,et al.Long-term effects of lanthanum intake on the neurobehavioral development of the rat[J]. Neurotoxicol Teratol,2006,28(1):119-124.
[4]He X,Zhang Z,Zhang H,Zhao Y,Chai Z.Neurotoxicological evaluation of long-term lanthanum chloride exposure in rats[J].Toxicol Sci,2008,103(2):354-361.
[5]Dong J,Yin H,Liu W,Wang P,Jiang Y,Chen J.Congenital iodine deficiency and hypothyroidism impair LTP and decrease C-fos and C-jun expression in rat hippocampus[J].Neurotoxicology,2005,26(3):417-426.
[6]杨敬华 ,蔡 原,刘秋芳,张立丰,齐 鸣,巫生文,等.镧影响大鼠学习记忆及海马c-fos基因、CA1区c-Fos蛋白表达的研究[J].毒理学杂志,2009,23(1):15-18.
[7]Mizuno M,Yamada K,Maekawa N,Saito K,Seishima M,Nabeshima T.CREB phosphorylation as a molecular marker of memory processing in the hippocampus for spatial learning[J].Behav Brain Res,2002,133(2):135-141.
[8]Ho N,Liauw JA,Blaeser F,Wei F,Hanissian S,Muglia LM,et al.Impaired synaptic plasticity and cAMPresponse element-binding protein activation in Ca2+/calmodulindependent protein kinase typeⅣ/Gr-deficient mice[J].J Neurosci,2000,20(17):6459-6472.
[9]Gélinas Y,Lafond J,Schmit JP.Multielemental analysis of human fetal tissues using inductively coupled plasma-mass spectrometry[J].Biol Trace Elem Res,1997,59(1-3):63-74.
[10]黄号栋,杨 静,龚 明.用磷酸二酯酶定量检测植物钙调素方法的改进[J].植物生理学通讯,2003,39(2):156-160.
[11]陈祖义.稀土元素的脑部蓄积性、毒性及其对人群健康的潜在危害[J].农村生态环境,2005,21(4):72-73,80.
[12]Feng L,Xiao H,He X,Li Z,Li F,Liu N,et al.Neurotoxicological consequence of long-term exposure to lanthanum[J].Toxicol Lett,2006,165(2):112-120.
[13]任丽宝,王小燕,解 清,刘景秀,黄 卓,杜 莹,等.大鼠脑组织中轻稀土元素含量相关性研究[J].分析试验室,2007,26(7):108-111.
[14]Lee HW,Yang W,Ye Y,Liu ZR,Glushka J,Yang JJ.Isolated EF-loopⅢ of calmodulin in a scaffold protein remains unpaired in solution using pulsed-field-gradient NMR spectroscopy[J].Biochim Biophys Acta,2002,1598(1-2):80-87.
[15]Wei F,Xia XM,Tang J,Ao H,Ko S,Liauw J,et al.Calmodulin regulates synaptic plasticity in the anterior cingulate cortex and behavioral responses:a microelectroporation study in adult rodents[J].J Neurosci,2003,23(23):8402-8409.
[16]Ye Y,Lee HW,Yang W,Shealy S,Yang JJ.Probingsite-specific calmodulin calcium and lanthanide affinity by grafting[J].J Am Chem Soc,2005,127(11):3743-3750.
[17]Hu J,Yang X,Wang K.La3+stimulate the activity of calcineurin in two different ways[J].J Biol Inorg Chem,2005,10(6):704-711.
[18]Lepsík M,Field MJ.Binding of calcium and other metal ions to the EF-hand loops of calmodulin studied by quantum chemical calculations and molecular dynamics simulations[J].J Phys Chem B,2007,111(33):10012-10022.
[19]Tardito D,Tiraboschi E,Kasahara J,Racagni G,Popoli M.Reduced CREB phosphorylation after chronic lithium treatment is associated with down-regulation of CaM kinaseⅣ in rat hippocampus[J].Int J Neuropsychopharmacol,2007,10(4):491-496.
[20]Barco A,Alarcon JM,Kandel ER.Expression of constitutively active CREB protein facilitates the late phase of long-term potentiation by enhancing synaptic capture[J].Cell,2002,108(5):689-703.
[21]Miyamoto E.Molecular mechanism of neuronal plasticity:induction and maintenance of long-term potentiation in the hippocampus[J].J Pharmacol Sci,2006,100(5):433-442.
[22]Guzowski JF.Insights into immediate-early gene function in hippocampal memory consolidation using antisense oligonucleotide and fluorescent imaging approaches[J].Hippocampus,2002,12(1):86-104.
