日本蟳捕食栉孔扇贝机制的初步研究
2010-09-24于宗赫杨红生刘保忠张立斌
于宗赫, 杨红生, 刘保忠, 邢 坤, 许 强, 张立斌
(1. 中国科学院 海洋研究所, 山东青岛 266071; 2. 中国科学院 研究生院, 北京 100049)
日本蟳捕食栉孔扇贝机制的初步研究
于宗赫1,2, 杨红生1, 刘保忠1, 邢 坤1,2, 许 强1, 张立斌1,2
(1. 中国科学院 海洋研究所, 山东青岛 266071; 2. 中国科学院 研究生院, 北京 100049)
在现场和室内条件下研究了日本蟳(Charybdis japonica)对栉孔扇贝(Chlamys farreri)的捕食机制。现场实验表明, 成年日本蟳可以捕食壳高小于5.0 cm的栉孔扇贝, 其捕食强度随着水温的升高而增大, 而壳高大于5.9 cm的栉孔扇贝则可以免遭日本蟳的捕食; 相对于栉孔扇贝, 日本蟳更倾向于捕食贻贝; 室内实验表明水温低于 10 ℃时, 日本蟳对大规格扇贝的捕食作用不明显。相同温度条件下,室内实验日本蟳的捕食强度要低于现场, 但其温度系数(Q10)差别不大。在上述实验基础上, 本文提出了提高底播栉孔扇贝成活率的方法, 即选择大规格的扇贝在水温较低的秋、冬季进行底播, 同时添加一部分贻贝作为牺牲诱饵以减少日本蟳对扇贝的捕食。
日本蟳(Charybdis japonica); 栉孔扇贝(Chlamys farreri); 捕食; 水温
海州湾前三岛海域刺参资源十分丰富, 近年来养殖业户为了获得更高的经济效益, 不断地在海岛周边底播刺参。刺参主要利用沉积物中的颗粒有机物为食, 然而, 自然沉积的颗粒物毕竟有限, 当刺参密度超过其养殖容量时就会导致该产业的滑坡。研究表明, 刺参可以有效地利用栉孔扇贝(Chlamys farreri)的生物沉积物作为食物来源[1,2], 因此, 在养殖刺参的海区底播扇贝, 一方面可以充分地利用海域资源, 另一方面, 底播扇贝可以通过“下行效应”加快水体中的颗粒物向底部转移, 增加海区刺参的食物量, 提高其养殖容量, 因此, 在前三岛海域进行栉孔扇贝底播其生态和经济效益都将非常可观。
扇贝底播养殖相对于筏式养殖来说可以节省大量的人力、物力, 然而底播养殖必须综合考虑海况、敌害以及死亡率等多方面的因素。自然条件下, 栉孔扇贝以足丝附着于海底营底栖生活, 前三岛海域海底多为岩礁和石块, 十分适合栉孔扇贝附着栖息。2007年夏初相关单位曾于前三岛海区进行过小规模栉孔扇贝(壳高 3~4 cm)的底播实验, 但是不久后调查发现底播扇贝成活率极低, 其中敌害生物的捕食是底播失败的主要原因。历次调查结果表明, 该海域日本 (Charybdis japonica)分布十分广泛, 并且生物量很大(约 30 g/m2), 在该海区进行栉孔扇贝底播,必须对这种生物的捕食作用进行充分考虑。
相关的研究表明, 敌害生物的捕食作用主要受水温以及底播贝类的规格等影响[3~6], 因此, 只有在适宜的水温条件下选择合适规格的扇贝进行底播才能获得成功。本研究在现场和室内条件下考察水温和扇贝规格对日本蟳捕食强度的影响, 以期为该海区进行栉孔扇贝底播生产提供科学依据。
1 材料与方法
1.1 现场实验
1.1.1 实验材料
栉孔扇贝, 取自前三岛海域筏式养殖区, 按照壳高分大、小两种规格, 其中: 大规格扇贝(B)壳高5.9~6.3 cm, 小规格扇贝(S)壳高 4.8~5.0 cm; 贻贝(Mytilus edulis) (M), 取自平岛周围岩礁区, 壳长2.9~3.3 cm。
日本蟳: 取自前三岛周围海域, 为附肢完整的健康成年雄蟹, 头胸甲宽为8.1~8.7 cm, 实验前暂养3~5 d, 期间不投喂食物。
1.1.2 实验设备
利用普通蟹笼进行诱捕和捕食实验, 蟹笼:Ø=40 cm,H=20 cm, 网孔1 cm, 共有4个入口, 根据实验需要开启或者封闭。
1.1.3 现场实验设计
于2007年11、12月初(该阶段水温较低、扇贝规格较大(壳高>4.5 cm)在海州湾平岛近岸海区进行,包括敌害生物诱捕实验以及捕食实验两种。判断日本蟳捕食的标准是: 栉孔扇贝(贻贝)有一壳破碎, 或者两壳都已破碎。
(1)敌害生物诱捕实验
于2007年11月初进行, 将不同规格的扇贝、贻贝进行搭配, 置于蟹笼中, 扇贝、贻贝搭配方式如下所示:
a.B: 10 粒/笼; b.S: 10 粒/笼; c.S+M: (10+10)粒/笼
上述3种搭配方式各设5平行组, 另外每种搭配方式各设立3个对照(蟹笼入口封闭), 考察实验期间扇贝(贻贝)自然死亡及笼外敌害生物透过笼壁对实验贝的捕食情况。将放置实验贝的蟹笼随机投放于实验海区, 笼间距约为2 m, 48 h后回收蟹笼观察敌害生物诱捕情况。
(2)日本蟳捕食实验
将蟹笼入口封住, 防止外部敌害生物进入, 11月份扇贝、贻贝规格及数目设置同诱捕实验a、b、c, 12月仅作日本蟳对扇贝捕食实验, 扇贝规格、数目设置同a、b, 各种搭配都设5个实验组以及3个对照组。上述实验组每笼放置日本蟳 1 只, 然后将蟹笼投放于实验海区, 48h后回收并记录各笼中被捕食扇贝数目。
1.1.4 环境因子监测
在实验开始和结束时利用多参数水质测定仪(YSI 6920)对实验海区环境因子进行监测, 重点考察不同实验期水温、pH、盐度以及溶解氧的变化。
1.2 室内模拟实验
栉孔扇贝(壳高 4.7~5.0cm)与日本蟳(雄性; 头胸甲宽 8.2~8.9 cm)均购自青岛市南山水产品批发市场。
利用50 cm×50 cm×50 cm PVC水缸进行实验,缸底随机投放一定数量的石块, 用于日本蟳隐蔽。日本蟳放于水缸后, 持续充气并利用加热棒对海水进行加热, 每天升温2~3 ℃, 达到预定温度后稳定3 d,然后每缸投放10粒栉孔扇贝, 每天早、晚各观察一次日本蟳对扇贝的捕食情况, 并及时补充扇贝。每个温度处理设1个对照缸, 即相同水温条件下, 仅投加栉孔扇贝, 观察不同实验温度条件下扇贝的自然死亡情况。室内实验温度梯度及实验结果设置见图1。

图1 日本蟳对扇贝、贻贝捕食结果Fig. 1 The predation rates of scallopChlamys farrerior blue musselMytilus edulisby crabCharybdis japonicain November and December 2007
1.3 温度系数
本研究将温度系数应用于捕食实验, 以量化水温对日本蟳捕食强度的影响。温度系数(Q10)计算公式为:
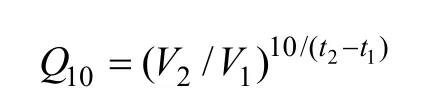
Q10即温度系数, 表示温度每升高 10℃, 捕食强度增加的倍数;V1、V2为捕食强度;t1、t2为相对温度。
2 实验结果
2.1 现场实验结果
2.1.1 环境因子监测结果
实验期间海区环境因子比较稳定, 两次实验期间海区盐度都为 30, pH为 8.3, 11月初平均水温为18.4℃, 溶解氧为 8.3 mg/L, 12月初平均水温为13.9℃, 溶解氧为9.2 mg/L。
2.1.2 敌害生物诱捕实验结果
2007年11月初海区敌害生物诱捕实验结果如表1所示, 三种搭配方式诱捕的敌害生物主要是日本蟳,其头胸甲宽为 6.9~8.1cm, 并且诱捕雌蟹数目要多于雄蟹。其中大规格扇贝组未捕获日本蟳; 小规格扇贝组共捕获 3只日本蟳, 其对扇贝的捕食数目为 1~6粒, 平均为 3.3粒; 小规格扇贝、贻贝混合组仅有一笼捕获日本蟳, 并且数目较多, 日本蟳对扇贝和贻贝都有不同程度的捕食。结果显示, 大规格扇贝对日本蟳的吸引力要小于小规格扇贝和贻贝, 但是不同的诱捕实验结果可能是蟹笼投放地点的差异造成的。
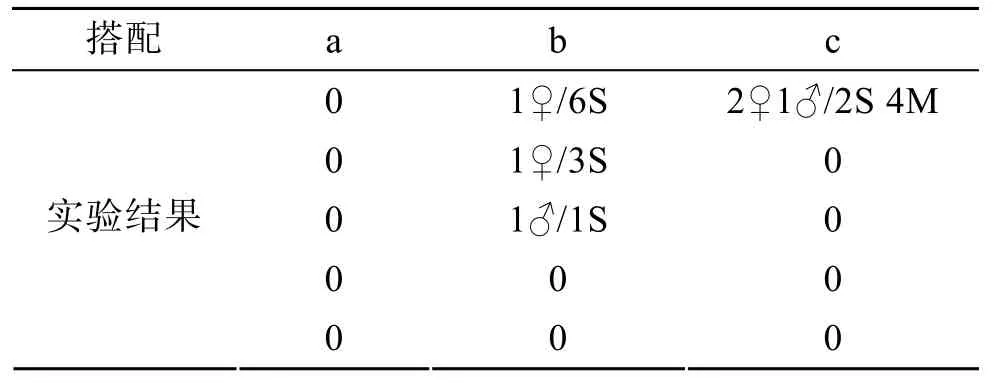
表1 2007年11月初敌害生物48h诱捕实验结果Tab. 1 Results of the predator entrapment in 48h in November 2007
日本蟳对扇贝以及贻贝的捕食实验结果如图 1所示, 可见这两个月份成年日本蟳对大规格扇贝(壳高>5.9cm)皆不构成危害, 并且随着水温降低日本蟳对于小规格扇贝的捕食强度下降; 日本蟳对于小规格扇贝与贻贝具有很强的捕食作用, 且更倾向于选择捕食贻贝。温度可以明显地影响日本蟳的捕食强度, 当水温降低时, 其捕食强度也大大降低。11月份个体日本蟳对小规格扇贝的捕食强度平均为1.9粒/d,12月平均为 0.6粒/d, 其温度系数Q10=13.0 (13.9~18.4 ℃)。
2.2 室内模拟实验结果
室内实验水温范围为 6~18℃(当温度超过 20℃时, 由于栉孔扇贝排放配子污染水质, 因此本研究没有涉及), 共设 4个温度梯度, 所用扇贝规格类似于现场实验所用的小扇贝。由图2可见, 在水温低于10℃情况下, 日本蟳对栉孔扇贝的摄食作用不明显,而当水温超过 12℃时, 日本蟳对扇贝的捕食强度随着水温的增加而提高很快, 其温度系数Q10= 15.6(12~18℃), 该值要略高于现场实验; 在相似水温条件下(现场 11月与室内 16~18℃; 现场 12月与室内12~14℃), 日本蟳现场实验捕食强度要高于室内实验, 其捕食率约为室内实验的3倍。

图2 不同温度条件下日本蟳对栉孔扇贝的捕食Fig. 2 The predation rates of scallopChlamys farreriby crabCharybdis japonicaat different water temperatures
3 讨论
日本蟳对栉孔扇贝的捕食强度受扇贝规格以及水温的影响非常明显: 现场条件下, 壳高超过5.9cm的栉孔扇贝可以免受日本蟳的捕食, 而壳高小于 5.0cm 的扇贝则极易遭受日本蟳的破坏; 水温在 10℃以下, 日本蟳对扇贝的捕食作用不明显, 而当水温超过14℃时,日本蟳的捕食作用随着水温的提高而迅速增加; 室内条件下日本蟳对扇贝的捕食强度不如现场实验, 而其温度系数Q10要略高于现场实验。
贝类规格、密度、水温、底质以及海流条件等都是影响敌害捕食率的重要因素[5,7,8]。自然条件下扇贝通常是采取闭壳而不是游泳的方式抵御蟹类的捕食, 因此非常容易被螃蟹捕获[9], 底播扇贝规格可以显著地影响蟹类的捕食强度[6,10,11]。从能量收支角度来说, 蟹类在单次捕食过程中, 选择大规格贝能量收益较高, 但是在处理猎物过程中体力消耗较大,并且存在捕食不成功的风险, 能量效率较低; 选择小规格贝则容易处理, 能量效率相对较高, 因此它们更倾向于捕食小规格的扇贝[12]。对于栉孔扇贝, 只有当其达到一定规格后, 贝壳才有足够强度抵御日本蟳的侵害, 因此, 选择较大规格的扇贝进行底播才能提高成活率。
水温对海洋无脊椎动物的代谢、运动速度影响很大[3,4], 它可以通过影响敌害生物的捕食及扇贝的应激反应速度决定底播扇贝的成活及在海底的分布[9]。相关的研究表明, 水温是影响蟹类捕食强度的重要因素[5], 对底播海湾扇贝的跟踪调查发现, 晚春和夏季水温高的时候, 底播区海青蟹(Callinectes sapidus)密度较大, 对扇贝的捕食率达到 100%, 而水温低的晚秋和冬季捕食作用几乎可以忽略[13]; 黄道蟹(Cancer irroratus)在较高水温条件下行动速度和反应速度都会提高, 其对巨扇贝的捕食作用也会明显提高, 其温度系数Q10=2.1[9]。本研究表明, 在较高水温条件下日本蟳对栉孔扇贝的捕食强度会大大提高, 这与上述研究结果基本一致。
相似水温条件下现场和室内实验日本蟳对栉孔扇贝的捕食强度差异较大, 这可能是由于实验条件的差异造成的。室内模拟条件不能完全反映现场情况, 因为水深、流速、底质情况等都无法模拟现场,相关的研究表明底质、海流等都可以影响蟹类的捕食, 如底质条件可能影响蟹类的行为, 同时也会对扇贝的活动造成影响[6]; 另外, 由于蟹类除了利用视觉系统寻找猎物外, 也可利用位于触角和步足之间的嗅觉系统来对猎物进行定位, 而海流能够影响水体中化学信号的传播, 因此相应地会影响捕食者对猎物的捕食[8], 本研究现场与室内实验结果的差异与海流可能也有一定关系。
室内实验所用日本蟳由于运输、暂养期间的损耗以及反季节加温等, 其活力会受到较大影响, 并且室内实验水缸由于较浅的水深, 日本蟳容易受到外界干扰, 所以捕食率相对于现场要低。
4 结论与建议
影响底播扇贝存活的主要因素是敌害生物捕食,在生产过程中, 养殖者可以通过控制多种因子来保证较高的成活率, 如底播密度, 扇贝规格以及底质类型等。选择大规格的扇贝进行底播会提高成功率, 然而这样势必延长中间养成时间, 增加养殖成本; 水温等环境因子也可以影响敌害生物的捕食以及贝类自身的存活。因此, 必须选择适宜的扇贝规格及时机进行底播,这样才能提高回捕率, 获得较高的经济价值。
研究表明, 前三岛海域深水区栉孔扇贝成活率要显著高于浅水区, 特别是在高温季节, 降低筏式养殖所在水层或者将扇贝养殖于底播设施之内可在很大程度上提高成活率[14,15], 而将扇贝直接进行底播则可大大降低养殖成本。该海区日本蟳生物量较大, 而日本蟳对扇贝具有较强的捕食作用, 其捕食是影响扇贝底播成功的主要因素之一, 由于日本蟳的捕食强度与水温以及扇贝规格密切相关, 因此应该从多方面入手以提高底播扇贝的成活率: 在苗种选择上尽量选择大规格扇贝进行底播; 底播扇贝的最佳时机应为水温低于10℃的秋、冬季以降低日本蟳的捕食作用; 底播前利用蟹笼等对海区的日本蟳进行诱捕; 日本蟳对小规格贻贝的捕食强度要大于栉孔扇贝, 因此, 在底播扇贝的同时掺杂一部分的贻贝作为“牺牲诱饵”, 也可以在很大程度上降低日本蟳对扇贝的危害, 前三岛周围海域岩礁区分布着丰富的贻贝资源, 这为该方法的实施提供了一定的基础。
[1] Zhou Y, Yang H S, Liu S L,et al. Feeding and growth on bivalve biodeposits by the deposit feederStichopus japonicusSelenka (Echinodermata: Holothuroidea)co-cultured in lantern nets [J]. Aquaculture, 2006, 256:510-520.
[2] 袁秀堂, 杨红生, 周毅, 等. 刺参对浅海筏式贝类养殖系统的修复潜力[J]. 应用生态学报, 2008, 19(4):866-872.
[3] Newell R C, Branch G M. The influence of temperature on the maintenance of metabolic energy balance in marine invertebrates [J]. Advances in Marine Biology,1980, 17: 329-396
[4] Cossins A R, Bowler K. Temperature biology of animals [M]. London : Chapman and Hall,1987.
[5] Nickell T D, Moore P G. The behavioural ecology of epibenthic scavenging invertebrates in the Clyde Sea area: laboratory experiments on attractions to bait in moving water, underwater TV observation in situ and general conclusions[J]. Journal of Experimental Marine Biology and Ecology, 1992, 159: 15-35.
[6] Wong M C, Barbeau M A. Effects of substrate on interactions between juvenile sea scallops (Placopecten magellanicusGmelin) and predatory sea stars (Asterias vulgarisVerrill) and rock crabs (Cancer irroratusSay)[J]. Journal of Experimental Marine Biology and Ecology, 2003, 287: 155-178
[7] Boulding E G, Hay T K. Crab response to prey density can result in density-dependent mortality of clams[J].Canadian Journal of Fisheries and Aquatic Sciences,1984, 41: 521-525.
[8] Powers S P, Kittinger J N. Hydrodynamic mediation of predator–prey interactions: differential patterns of prey susceptibility and predator success explained by variation in water flow[J]. Journal of Experimental Marine Biology and Ecology, 2002, 273: 171-187.
[9] Barbeau M A, Scheibling R E. Temperature effects on predation of juvenile sea scallops (Placopecten magellanicus(Gmelin)) by sea stars (Asterias vulgarisVerrill)and crabs (Cancer irroratusSay) [J]. Journal of Experimental Marine Biology and Ecology, 1994, 182:27-47.
[10] Kamenos N A, Moore P G, Hall-Spencer J M. Maerl grounds provide both refuge and high growth potential for juvenile queen scallops (Aequipecten opercularisL.)[J]. Journal of Experimental Marine Biology and Ecology, 2004, 313:241-54.
[11] Barbeau M A, Scheibling R E. Behavioral mechanisms of prey size selection by sea stars (Asterias vulgaris Verrill) and crabs (Cancer irroratusSay) preying on juvenile sea scallops (Placopecten magellanicus(Gmelin)) [J]. Journal of Experimental Marine Biology and Ecology, 1994, 180: 103-136.
[12] Juanes F, Hartwick E B. Prey size selection in dungeness crabs: the effect of claw damage[J]. Ecology, 1990,71: 744-758.
[13] Bishop M J, Rivera J A, Irlandi E A,et al. Spatio-temporal patterns in the mortality of bay scallop recruits in North Carolina: investigation of a life history anomaly[J]. Journal of Experimental Marine Biology and Ecology, 2005, 315(2): 127-146.
[14] Yu Z H, Yang H S, Liu B Z,et al. Growth, survival and immune activity of scallops,Chlamys farreriJones et Preston, compared between suspended and bottom culture in Haizhou Bay, China[J]. Aquaculture Research,2010, 41: 814-827.
[15] Yu Z H, Liu B Z, Yang H S,et al. Survival, growth, and immune activity of scallopChlamys farrericultured at different depths in Haizhou Bay during the warmest months of the year[J]. Chinese Journal of Oceanology and Limnology, 2010, 28(3): 498-507.
Received: Jun., 10, 2009
Key words:Charybdis japonica;Chlamys farreri; predation; water temperature
Abstract:Predation of scallopChlamys farreriby crabCharybdis japonicawas examinedin situand in the laboratory at different water temperatures. It was found that the predation rate of small scallop (shell height < 5.0 cm)was increased with increasing water temperature, and the predation was negligible either when the shell height of scallop exceeded 5.9 cm or when the water temperature was blow 10°C.In situpredation rate was higher than that in the laboratory at the same seawater temperature; however, theQ10values were similar at the two different situations. Our results indicate that the best way to enhance the survival of the bottom culturedC. farreriis to seed large scallops in autumn and winter, when the water temperature is low, and some mussels should also be put in the culture area as a sacrifice to reduce the predation of scallop by crab.
(本文编辑:张培新)
Predation of scallop Chlamys farreri by crab Charybdis japonica
YU Zong-he1,2, YANG Hong-sheng1, LIU Bao-zhong1, XING Kun1,2, XU Qiang1,ZANG Li-bin1,2
(1. Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Graduate University, Chinese Academy of Sciences, Beijing 100049, China)
S944.4+3
A
1000-3096(2010)12-0062-05
2009-06-10;
2009-08-03
国家科技支撑计划项目(2006BAD09A02); 国家863计划项目(2006AA100304)
于宗赫(1980-), 男, 山东青岛人, 博士研究生, 从事养殖生态学研究, 电话: 0532-82898705, E-mail: ouqdyuzh@yahoo.com.cn; 杨红生, 通信作者, 研究员, 电话 0532-82898620, E-mail: hshyang@ms.qdio.ac.cn
